
04/06/2002 11:29 PM
Triassic Strata of the Colorado Plateau
Henry Morris wrote:
"Now the geologic column is an idea, not an actual series of rock layers. Nowhere do we find the complete sequence. Even the walls of the Grand Canyon included only five of the twelve major systems (one, five, six and seven, with small portions here and there of the fourth system, the Devonian." (Morris and Parker, 1987, p. 163)
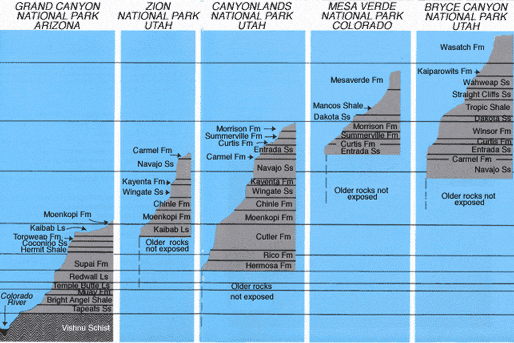
Morris neglects to mention that, to the north and east of the Grand Canyon, the Kaibab is in fact overlain by Triassic, Jurassic, Cretaceous and Cenozoic deposits. When strata of the Grand Canyon are considered together with the overlying strata of the Grand Staircase, we have an approximately 2 mile thick column of sedimentary rocks, in which 10 of the 12 periods (Ordovician and Silurian not present) are represented by sediments, rather than the 5 suggested by Morris and Parker. Contrary to Morris' statement, moreover, many intracratonic basins do in fact contain deposits from each of the 12 phanerozoic systems. At least two dozen such basins are known (Robertson Group, 1989. Stratigraphic Database of Major Sedimentary Basins of the World, Llandudno Gwynedd, England). See, for example, Glenn Morton's online article The Entire Geologic Column in North Dakota for a detailed description of the geologic column as it exists in the Williston Basin.
The Grand Canyon actually cuts through part of the southern slope of a north-south trending bulge in the earth, the Kaibab uplift. This bulge is a block of earth that has been "pushed up" thousands of feet compared to surrounding areas. This is why, for example, the north rim of the canyon is about 1000ft higher than the south rim. This uplift greatly accelerated denudation in the area of the Grand Canyon, but did not affect areas surrounding the uplift. Thus, as one traces the Kaibab Formation to the east and north of the Kaibab uplift, a series of ascending cliffs and terraces come into view. This series of cliffs and terraces, composed of Triassic and younger strata, is referred to as the Grand Staircase. The total thickness of these additional strata, about 1 mile, is roughly equal to that of the strata in the Grand Canyon.
Mesozoic and Cenozoic Strata of the Grand Staircase: North - South Section
[vertical scale exaggerated]
There is abundant evidence that massive amounts of sedimentary rock have been denuded from the Grand Canyon area. Baar writes that "although the Kaibab Formation is the uppermost rock unit that is evident along the Grand Canyon, it was not the last event in the history of the region. At least ten thousand feet of younger rocks were deposited above the Kaibab in Mesozoic time, as shown by the remnants of these rocks surrounding the Kaibab uplift or monocline" (p. 29). Morales notes similarly that, although the Mesozoic and Cenozoic deposits have been denuded in two major episodes (one in the late Cretaceous and another in the Oligocene) as a result of uplifting in the southwestern Colorado Plateau, "[w]idely dispersed remnants of these deposits . . . still crop out in the Grand Canyon area" (p. 248). For instance, a small outcrop of Moenkopi and Chinle sediments is present at Ceder Mt in the southeast corner of the Grand Canyon park. In addition, there are caverns in the Redwall Limestone which often contain remnant clasts of these previously existing, overlying sedimentary rocks, including clasts from the Chinle Formation (Wenrich, K.J. and Hunton, P.J. Breccia Pipes and Associated mineralization in the Grand Canyon Region, Northern Arizona, Geology of the Grand Canyon, Northern Arizona, 28th Int. Geol. Congress, Field Trip Guide Book, Washington: AGU, 1989, p. 212). Since the next outcrop of the these sediments is 95 miles to the north of this area, this shows that at least 1000ft or so of these sediments were denuded from the Grand Canyon area.
The Moenkopi Formation (Triassic)
As noted previously, an erosion surface is present at the Kaibab-Moenkopi contact in northwestern Arizona, southeastern Nevada, and southwestern Utah, including channels 250ft deep and 700ft wide in Rock Canyon, and possible paleokarst depressions. The Moenkoi Formation, which is about 1,800 ft at its thickest, is composed of fluvial (stream) deposits of silt, sand and mud in the east, which grade westwardly into marine limestone. In areas where large paleostreams can be reconstructed, they appear to have flowed to the west and to the north. Baar writes:
"The formation, generally speaking, changes most emphatically from east to west in a definite pattern. In the eastern Colorado Plateau of southwestern Colorado and northwestern New Mexico, the Moenkopi is either missing or is thin and composed of deposits that suggests largely a fluviatile origin. As one progresses westward, the formation gradually thickens and contains increasing amounts of mudstones and shales, which are hirizontally bedded with extensive development of symmetrical and current ripple marks, mud cracks, and other surface features of the bedding planes that require shallow, tidal-flat conditions to form.
"West of the intertidal facies the Moenkopi gradually displays more and more marine characteristics as it thickens to a maximum of two-thousand feet. Marine limestones become prominent until they form a major part of the formation along the western margin of the Colorado Plateau . . . Still farther west and northwest, the Moenkopi grades into a totally marine sections known as the Thaynes Limestone, which contains ammonitic cephalopods of Lower Triassic age" (p. 158-160).
Raindrop prints and salt casts are present on some bedding planes also (Chronic, 1990, p. 195). The bulk of the Moenkopi sediments appear to have been deposited by streams moving west-northwest from a cratonic area of low relief (Lemon, 1993, p. 324).
Both subaqeous and subarial tracks and traces are known from the Moenkopi, from over a dozen stratigraphic levels. Terrestrial or subaerial tracks and traces (ichnotaxa) in the Moenkopi include the vertebrate traces Chirotherium, Rhynchosauroides, Synaptichnium, Therapsipus, and Rotodactylus. Five taxa of invertebrate (subaerial and subaqueous) trace fossils are found in the Moenkopi and include subaqueous arthropod trails made by limulids (Peabody, 1956; Morales, 1987; Hunt et al., 1993).
Note that a virtually identical vertebrate footprint fauna is present in middle Triassic deposits of the Central Massif of France. Taxa recognized here include 'Anchisauripus' [possibly representing an early, Coelophysis-type dinosaur], Chirotherium, Brachychirotherium, Rhynchosauroides, Synaptichnium, and Rotodactylus. See Demathieu, G.R. Trace Fossil Assemblages in Middle Triassic Marginal Marine Deposits, Eastern Border of the Massif Central, France, in Biogenic Structures: Their Use in Interpreting Depositional Environments. Curran, H.A. ed. SEPM Special Publication No. 35, 1985.
The Chinle Formation (Triassic)

Paleogeographic reconstruction of the Colorado Plateau during early Chinle time. From Ron Blakey's Stratigraphy and Sedimentology on the Colorado Plateau website.
The Chinle Formation consists of a conglomerate valley-fill sequence overlain by fluvial, floodplain, marsh, delta, and lacustrine deposits (Stewart et al., 1972; Dubiel et al., 1991). The depositional environment inferred for the Chinle Formation is similar in many respects to the modern Congo Basin. Information on the Congo Basin can be found here, here, and here. Note that the Congo Basin is actually about 30% larger than the Chinle Basin would have been.
"The Upper Triassic Chinle Group encompasses strata assigned to about 50 lithostratigraphic units in the western United States ranging from West Texas to Nevada and Idaho to New Mexico. Most of these strata were deposited in a single depositional basin associated with a large fluvial system with paleoflow to the west and a drainage basin comparable to the Congo Basin of equatorial Africa. The depositional environments include channels, overbank/crevasse splays, floodplains, and minor lacustrine facies, with increasing evidence of eolian and lacustrine deposition at the top of the section. Three unconformity-bound sequences have been observed in these deposits. The first two sequences consist of basal channel-fill conglomerates and sandstones that fine upwards into dominantly flood-plain mudstones and paleosols. The third sequence consists primarily of interbedded siltstones and fine sandstones, and demonstrates an increasing eolian trend."
The basal Chinle member, the Shinarump Conglomerate, consists of conglomerate-filled channels up to 40 meters thick. In some areas the Shinarump Conglomerate contains clasts as large as a car. The conglomerate also contains gravelly fossiliferous clasts derived from the underlying Kaibab limestone. Above this conglomerate lie fluvial sediments of mud and silt. The upper members are colorful shales with a few thin beds of fluvial sandstone. Bentonite volcanic ash horizons are present also. Outcrops of these two members constitute some of the most beautifully colored areas of the Colorado Plateau, or of the entire world for that matter.
Based on tetrapod track assemblages (summarized in Hunt and Lucas, 1995), plant fossils (Ash, 1980, 1987b), pollen content (Litwin et al., 1991), and calcareous microfossil content (Lucas and Kietzke, 1995), the Chinle is assigned a late Triassic (Norian) age, about 215-220mya (Lucas, S. G. 1993. The Chinle Group: revised stratigraphy and biochronology of Upper Triassic nonmarine strata in the western United States. Museum of Northern Arizona Bulletin, 59:27-50.). Two radiometric ages have been published for the the reworked tuff deposits found in the highest member of the Chinle, a K-Ar biotite date of 239±9 Ma, and a U-Pb zircon date of 207±2 Ma (Riggs, N. R., S. R. Ash, and J. M. Mattinson. 1994a. Isotopic dating of a non-volcanic continental sequence, Chinle Formation, Arizona. Geological Society of America Abstracts with Programs, 26(6):61).
The Chinle preserves a variety of fossils of plants and animals. Animal fossils include Metoposaurus, a metoposaurid amphibian, Postosuchus and Sphenosuchus, the aetosaurs Calyptosuchus and Typothorax, the phytosaur Pseudopalatus, and others. Over 70 plant taxa are known from the Chinle, including bennettitaleans, horsetails, the ferns Phlebopteris smithii, Cynepteris lasiophora, and Cladophlebis, the cycads Charmorgia, Eoginkgoites davidsonii, and Zamites powellii, the lycopods Chinlea and Seleginella anasazia, the conifers Araucarioxylon, Pagiophyllum sp., Brachyphyllum sp., Masculostrobus clathratus, Dechellyia gormanii, and cuticle of the conifers Brachyphyllum, Pagiophyllum, and Dinophyton (Pig, K.B. and Davis, W.C. Anatomically Preserved Plant Reproductive Structures from the Upper Triassic Chinle Formation in Petrified Forest National Park, Arizona. NPS Paleontological Research, Vol. 2, 177). The upper Black Forest Member includes not only Araucarioxylon, but also the less common genera Woodworthia and Schilderia. Many of these logs have been permineralized into agate or jasper. Most of the petrified logs seem to have been transported, but some in situ stumps have been reported (Ash and Creber, 1992. Palaeoclimatic interpretation of the wood structures of the trees in the Chinle Formation, Petrified Forest National Park, Arizona, USA. Palaeogeography, Palaeoclimatology, Palaeoecology, 96:299-317).
Heckert and Garza note that "existing biostratigraphic correlations in the Chinle Group allow the discrimination of at least four stratigraphically superposed Upper Triassic vertebrate faunas that span from Carnian to late-Norian (Rhaetian?) age. This is the best-studied succession of Late Triassic nonmarine vertebrate faunas in the world, and can be used to correlate Chinle strata to Triassic basins in eastern North America, Greenland, Europe, North and South Africa, South America, and India." The flood theory will need to explain this nonrandom, stratigraphic sorting of fossils within the Chinle itself, and the fact that very similar biostratigraphic arrangements are found in contemporaneous Carnian-Norian deposits elsewhere in the world.
Also preserved in the Chinle are a numer of trackways, including those of large amphibians, various archosaurs such as Anchisauripus sillimani and Rhynchosauroides, some early, small dinosaurs (such as Coelophysis [also called Rioarribasaurus] and some Prosauropods), and a few mammal-like reptiles. A very similar vertebrate fauna, both of fossils and footprints, is preserved in Triassic deposits of the Newark Supergroup on the east coast.
Whereas footprints of mammal-like reptiles are dominant in underlying Permian strata, in early Triassic strata small Archosaurian tracks (Chirotherium-like) appear and become the predominant land vertebrates. The most well-known members of the Archosauria include dinosaurs, pterosaurs, and crocodilians. This faunal displacement is evident in the Chinle Formation and the overlying Glen Canyon Group, in which three toed Grallator type tracks, attributed to early dinosaurs, become increasingly abundant, and increasingly large, as a function of stratigraphic height. This change in track type proportions is found globally in mid to late Triassic terrestrial deposits, and marks the beginning of a long dinosaurian dominance of terrestrial vertebrate faunas. Lockley and Hunt (1995) note:
"Toward the end of the Triassic, in youngest Chinle Group sediments, small-sized Grallator tracks appear in association with a diverse assemblage of other archosaur tracks. Then in some of the very youngest Chinle layers (for example, the Sheep Pen Sandstone) these Grallator tracks become the dominanat track type to the virtual exclusion of all other footprint types. We get the impression that the small dinosaurs were taking over. This impression if confirmed by the track record in the overlying strata of the Wingate Formation, where we find more medium-sized Grallator tracks, and very few other track types" (p. 103).
Chinle deposits include a variety of invertebrate burrows and tracks as well, including many attributed to horseshoe crabs, millipedes, adult and juvenile insect burrows and nests, beetle borings and gallery nests in wood, annelid burrows, and Scoyenia, a burrow attributed to insect larvae. Hasiotis et al. (Continental Trace Fossils, Petrified Forest National Park: Tools for Paleohydrologic and Paleoecosystem Reconstructions, Technical Report NPS/NRPO/NRTR-93/11) describe 6 types. Camborygma burrows with diameters in centimeters and lengths from 30 cm to 200+ cm are known from the upper Chinle, and in the correlative Dockum Formation of northwest Texas and eastern New Mexico. These may include chimney structures at their tops, and a "surficial burrow morphology including scrape marks, scratch marks, mud-and lag-liners, knobby and hummocky surfaces, pleopod striae, and body impressions" (Hastiostis et al.. nd). Hastiostis (1990; 1991) and Hasiotis and Mitchell (1993) attribute these to crayfish, which produce burrows with virtually identical features. Scoyenia burrows have a rope like shape, a diameter of 0.2 cm to 10 cm, and lengths from a few cm to 10 cm. These burrows are attibuted to insect larvae. Koupichnium tracks, attributed to freshwater horseshoe crabs are of two types "1) Two cheveron-like series of tracks each of 4 oval to round holes or bifid V-shaped impressions or scatches, and 2) one pair of digitate or flabellar, toe-shaped imprints with or without a medial drag mark." Cylindricum burrows are vertically oriented, possess round bottoms, and have a diameter up to 5 cm and lengths up to several centimeters. Archeoentomoichnos burrows are perhaps the most interesting. These consist of "multistory ramps, floors, and walls constructed in a cylindrical structure approximately 7 cm in diameter; associated with mm to cm diameter-sized corridors greater than 5 cm in length; walls, ramps, and floors, range in thickness from 2 mm to 5 mm. . . These nests represent the earliest known examples of social behavior in insects as well as the oldest evidence of termite activity. The ichnofossil evidence predates the body fossil evidence of termites by 135 million years (early Cretaceous). The nests are composed of calies (nest proper), galleries (runways between nests), and peripheral calies (storage chambers). Nests like these termite edifices are important because they preserve the behavior of a group of organisms that reflect the division of labor amount individuals that allow such colonies of insects to perpetuate themselves." These structures are considered by Hasiotis to be clear examples of hymenopteron nests.
Paleosols of varying degrees of maturity are found throughout the Chinle. Hasiotis et al. write:
"Major types of paleosols present in the Chinle include Gleysols, Alfisols, Vertisols, Calcisols, and Aridisols, all of which also range in stages of maturity based on the amount of time in their formation (e.g., Mack et al., 1993).
"Gleysols are abundant in the basal Chinle (Shinarump Member/"mottled strata"), where they are characterized by extensive purple, yellow, and white mottled horizons, contain deep crayfish burrows and roots, and indicate deep though fluctuating water tables.
"Alfisols are common in floodplain mudrocks in the lower and middle Chinle (Monitor Butte and Petrified Forest Members), consisting of thick red, clay-rich horizons, locally exhibiting small carbonate nodules. These soils contain red-purple mottles, abundant beetle burrows (Scoyenia) and small rhizoliths, and indicate predominantly moist soils and persistently high water tables.
"Vertisols (Monitor Butte and Petrified Forest Members) are characterized by clay-rich horizons, deep mudcracks, slickensides, gilgai micro-relief, carbonate nodules, crayfish burrows and extensive rhizoliths, and are indicative of periods of wetting and drying.
"Calcisols (Owl Rock and Church Rock Members) with carbonate accumulations in their upper portions, contain rhizotubules and rhizocretions, and occur predominantly in siltstone deposits.
"Aridisols (Church Rock Member) contain varying stages of carbonate nodule development, few rhizoliths, rare bioturbation, and indicate decreased precipitation coupled with persistently deeper water tables" (A Holistic Approach to Reconstructing Triassic Paleoecosystems: Using Ichnofossils and Paleosols as a Basic Framework, National Park Service Technical Report NPS/NRGRD/GRDTR-98/01).
See also: National Park Service Paleontology Program, Petrified Forest National Park, Trees to Stone.
The Triassic-Jurassic Extinction At the Triassic -Jurassic boundary, many of the prominent Triassic non-dinosaurian trackmakers disappear. Globally. These include Rhynchosauroides, Tetrasauropus, Brachychirotherium, Atreipus, and others. Dinosaur tracks dominate thereafter, globally, until the end of the Cretaceous. This change in footprint fauna is one manifestation of an "extinction-event" which dramatically affected both marine and terrestrial organisms at or very near the Triassic-Jurassic boundary. Both impact craters and shocked quartz grains are known from the Triassic-Jurassic boundary, although it is not clear that impact provided the actual extinction mechanism. Lemon (1993, p . 333) writes:
Range Chart Showing a pronounced extinction at the Triassic-Jurassic Boundary. "Ranges of families of tetrapods through the Triassic and Early Jurassic depicted at the stage level. The darker blue bars ranges are from Olsen (1986) and the lighter blue bars are range extensions since that time. Note that the largest concentrations of extinctions occurs at the Triassic-Jurassic boundary." Whatever the boundary event, dinosaurs were barely if at all affected. From online article Triassic-Jurassic Boundary on the ldeo.columbia.edu server. You can find more info on the end-Triassic extinction here. |
