 Fig.1: Gravat de Robert Hooke on apareix l'estructura cel·lular
d'una làmina de suro
Fig.1: Gravat de Robert Hooke on apareix l'estructura cel·lular
d'una làmina de suro1. Introducció: de la cèl·lula procariota a la cèl·lula eucariota
1.1. Tipus de microscopis
1.2. Teoria cel·lular
1.3. L'origen de la vida
1.4. Nivells d'organització cel·lular
1.4.1. Característiques generals de les cèl·lules procariotes
1.4.2. Característiques generals de les cèl·lules eucariòtiques
1.4.3. Comparació entre les cèl·lules procariotes eucariotes.
1.4.4. Aspectes evolutius de la cèl·lula.
Abans d'encetar el tema us propose visitar aquesta WEB que us donarà una bona idea general d'aquest tema
1. Introducció: de la cèl·lula procariota a la cèl·lula eucariota
Fig.1: Gravat de Robert Hooke on apareix l'estructura cel·lular
d'una làmina de suro
 Fig.2: Microscopi de de Robert Hooke
Fig.2: Microscopi de de Robert Hooke
Tots els microscopis utilitzen radiacions amb les quals il·luminen la mostra. La capacitat d'ampliació d'un microscopi serà tant més gran quant menor siga la longitud d'ona de la radiació utilitzada. Per aquest motiu, es van inventar microscopis que usen electrons, que tenen una longitud d'ona molt més petita que la llum visible. Podem classificar els microscopis en dos grans grups
- Microscopis òptics
Aquests microscopis que usen llum visible. pot observar cèl·lules vives o mortes. Els materials observats mostren colors, bé sigui els seus naturals o els que han adquirit al sometre'ls a un procés de tinció. El microscopi òptic té un límit de resolució al voltant de 200 nm (0.2 µm ). aquest límit es deu a la longitud d'ona de la llum (0.4-0.7 µm ).

Fig.3: Cèl·lules epidèrmiques de ceba, tenyide si onservades al microscopi òptic. Teixit animal
Per poder observar mostres al m.o., aquestes han de ser suficientment primes per deixar passar la llum al seu través. El teixit cal fixar-se (amb alcohol, per exemple, per evitar la seua descomposició. Seguidament es talla en làmines de poques micres amb l'ajuda d'un instrument anomenat microtom. Normalment la mostra es tenyeix amb uns colorants específics que es fixen a determinades estructures cel·lular i permeten una millor visió
2. Microscopis electrònicsEls microscopis electrònics utilitzen feixos d'electrons. En aquest cas els materials no presenten cap coloració, només són més o menys opacs front als electrons, el que es veu com diferents tons de gris. Únicament es pot observar cèl·lules mortes. En distingirem de dos tipus:
- m.e. de transmissió (MET)
- m.e. d'escombrada o scanning (MES)
Components del microscopi òptic
Part òptica:

fig.4: Components del microscopi òptic.
Part mecànica:
Ací teniu un enllaç amb un centenar de fotografies de microscopia òptica comentades
Microscopi electrònic de transmissió
El microscopi electrònic de transmisión (MET) té un límit de resolució al voltant de 2 nm. Això és degut a limitacions de les lents usades per enfocar electrons cap a la mostra. Un MET mira a repliques de cèl·lules mortes , després d'haver estat fixades i tenyides per ions de metalls pesants. Els electrons són dispersats quan passen a través d'una fina secció del material a estudiar i posteriorment detectats i projectats cap a una imatge sobre una pantalla fluorescent.
![]()

Microscopi electrònic
de escombrada (scanning)
El microscopi elctrònic
dèscombrada (MES) ) també té un límit
de resolució al voltant de 2 nm. A l'igual que el MET,
el MES permet d'observar cèl·lules mortes , després
d'haver estat fixades i tenyides per ions de metalls pesants.
Amb aquesta tècnica els electrons són reflectits
sobre la superfície de la mostra.La imatge té un aspecte tridimensional
 Fig.5: Trajectòria dels
electrons en el MET i el MES
Fig.5: Trajectòria dels
electrons en el MET i el MES



Fig.6: Cèl·lules de la sang observades amb el MES
Si vols utilitzar un simulador d'un microscopi electrònic d'escombrada (scanning) ací tens la teua oportunitat

Fig.7: Mides relatives
de diferents tipus d'éssers vius i els seus components
Els anys 1838-39, el botànic
M. Schleiden i el zoòleg T. Schwann
van aportar les proves definitives, basades en les seues
investigacions i en les dades que hi havia acumulades, que van
permetre establir l'enunciat fonamental de la teoria cel·lular
: les plantes i els animals són constituïts per
una o més unitats fonamentals, les cèl·lules.
Uns anys després Virchow va completar aquest enunciat
quan va proclamar que cada cèl·lula procedeix d'una
altra cèl·lula "omnis cellula ex cellula"
L'enunciat actual de la teoria cel·lular es pot resumir
en tres punts:
1- La cèl·lula és la unitat anatòmica de tots els éssers vius, ja que tots estan constituïts per una o més cèl·lules (a excepció del virus)
2- La cèl·lula és la unitat de funcionament dels éssers vius: tota cèl·lula duu a terme les funcions bàsiques de tots els éssers vius: l'obtenció de matèria i energia (funcions de nutrició) i l'aplicació d'aquesta energia a autoperpetuar les seues estructures al llarg del temps (funcions de relació i reproducció).
3- La cèl·lula és la unitat d'origen dels éssers vius, és a dir, que cada cèl·lula procedeix d'una altra preexistent, per divisió d'aquesta.
Un dels postulats més importants de la biologia fou la demostració de la inexistència de la generació espontània: tots els éssers vius provenen d'un altre ésser viu preexistent, tal com ho va desmostrar Pasteur al segle XIX. Irònicament, avui en dia, sabem que l'inici de la vida es realitzà de forma espontània a partir de components moleculars abiòtics existents en el nostre planeta fa ara més de 3500 ma.
L'evolució abiòtica que originà la vida va seguir els passos següents:
Si volem entendre aquesta creació de la vida a partir de material abiòtic, cal considerar dues idees:
- L'extensió de la idea de la selecció natural a nivell de les biomolècules
- El fet que les condicions de la Terra primitiva eren molt diferents de les actuals: presència d'una atmosfera no-oxidant, ja que els nivells d'oxigen de l'actualitat són el resultat de més de 2000 ma de fotosíntesi i l'abundància de molècules orgàniques produïdes de forma abiòtica


Fig.8: Esquema de l'experiència de Miller i Urey. Fotografia de Stenley Miller al seu laboratori
Stanley Miller i Harold Urey utilitzaren un aparell semblant al de la figura per simular les condicions existents en l'atmosfera primitiva de la Terra. un recipient d'aigua calenta simularia el mar primitiu, mentre que l'atmosfera estaria constituída per H2O, H2, CH4, and NH3. Un corrent elèctric representaria les abundant tempestes elèctriques existenta aleshores. per últim un condensador refredaria l'atmosfera i crearia pluges. Després d'uns dies de funcionament l'aigua del recipient passava de ser transparent a adoptar un color marró on es podien detectar una varietat de molècules orgàniques entre les quals es trobaven els aminoàcids de formen les proteïnes. quedava demostrat que les molècules orgàniques que formen part dels éssers vius poden ser sintetitzades abiòticament d'una manera prou senzilla.
Entrevista a Stanley Miller en la revista MÈTODE de la Universitat de València
L'energia necessària per a l'aparició d'aquestes molècules podia procedir d'una llista variada de fonts:
1. Radiació còsmica o la procedent d'isòtops inestables de la Terra
2. Llum UV ja que aleshores no havia cap capa d'ozó.
3. Descàrregues elèctriques
4. Calor procedent de l'abundant activitat volcànica
Com va aparèixer la primera cèl·lula?
Aquesta història no és molt fàcil d'explicar. Pensem en la presència de polinucleòtids que tenien la capacitat de crear còpies de si mateixos gràcies a la complementarietat de les seues bases constituents. Els diferents polinucleòtids competien entre si per aconseguir els nucleòtids necessaris per poder replicar-se. aquesta replicació era lenta i plena d'errades. Poc a poc els nucleòtids eren més i més complexos. Hi havia enzims que catalitzaren aquestes reaccions? (encara que els ions metàl·lics existents en el mar primitiu podien haver actuat com a biocatalitzadors). Una polèmica molt estesa consisteix en preguntar-se "què va ser primer: les proteïnes (enzims) o els àcids nucleics"?. Les mutacions i l'evolució a nivell molecular feien que aquesta replicació fos més estable. De tota manera sembla ser que els primers polinucleòtids eren cadenes senzilles d'ARN. La vida actual, amb l'excepció d'alguns virus, té a l'ADN com a molècula portadora de la informació genètica. Una raó per pensar que l'ADN substituís l'ARN és que l'ADN, amb les seues dues cadenes, és una molècula més estable. En l'actualitat sabem que determinats ARNs tenen capacitat catalítica, es a dir, que a més de poder crear òpies, poden actuar com a catalitzadors de les reaccions metabòliques. No sabem a hores d'ara com s'establí la interrelació entre cadenes d'ADN i de proteïnes (enzims). Altre aspecte important en l'aparició de la primera cèl·lula (més senzilla encara que els actuals bacteris) va ser l'existència d'una membrana plasmàtica que independitzara l'ADN del medi ambient i facilitara la seua replicació. Ara sabem que les molècules de fosfolípids poden crear espontàniament bicapes sota determinades condicions. El resultat de tota aquesta evolució al llarg de milions d'anys va ser l'aparició del primer ésser autopoiètic, la primera cèl·lula.
Per a saber més sobre l'origen de la vida
Senzilla escala del temps del procés vital
Com vam veure al primer tema en
parlar dels nivells d'organització de la matèria
viva, el nivell cel·lular estava constituït per un
conjunt de molècules, les quals havien arribat a atènyer
un grau de complexitat suficient per manifestar propietats vitals.
Un organisme viu no és simplement un mosaic d'estructures
moleculars independents, sinó un sistema integrat i autoregulat,
en el qual estructura i funció són inseparables.
En estudiar les cèl·lules procedents de diferents
organismes o teixits, es pot observar un cert grau de diversitat
cel·lular, sobre tot pel que fa a la forma i la grandària,
tot i que totes presenten unes característiques bàsiques
comunes:
a) un embolcall que separa la cèl·lula de l'exterior i que regula l'intercanvi amb l'exterior
b) Un contingut cel·lular on es realitzen tots els processos químics (citoplasma).
c) Un material hereditari (ADN) que codifica tota la informació genètica.
Segons el grau de complexitat, però,
es poden considerar dos tipus d'organització cel·lular:
la cèl·lula procariota i la cèl·lula
eucariota. A més d'aquests dos tipus d'organització
cel·lular es considera un tercer tipus d'organització:
el que formen els virus, l'extraordinària simplicitat
dels quals, al costat de la seva mancança de metabolisme
propi, els converteix en organismes que se situen en la frontera
entre les estructures vives i les inertes.

Fig.9: Les cèl·lules procariotes formen el regne
dels monera, mentre que la cèl·lula eucariota (amb
algunes diferències) és la component de la resta
dels 5 regnes en què estan dividits els éssers vius:,
protista, plantae, fungi,i animals.
Els procariotes són el conjunt d'organismes unicel·lulars que constitueixen el regne dels monera, en el qual s'inclouen els bacteris, les algues cianofícies i els micoplasmes.
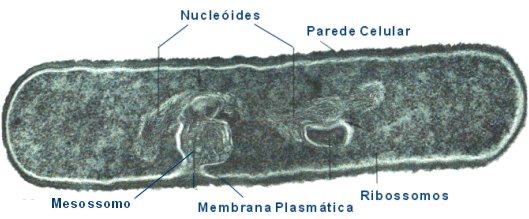
Fig.10: Cèl·lules
procariotes
La principal característica de la cèl·lula procariòtica es l'absència d'una membrana nuclear que aïlle el seu ADN de la resta de la cèl·lula. El seu material genètic està constituït per una única molècula d'ADN bicatenari, circular i nu (no associat amb histones). Són cèl·lules molt més menudes i d'estructura molt senzilla ja que manquen de la major part dels orgànuls típics de la cèl·lula eucariota. Parlarem breument de les principals característiques estructurals dels bacteris:
Elements estructurals dels bacteris
La càpsula bacteriana és una capa gelatinosa formada per polisacàrids ii proteïnes. Presenta dues funcions: regulació dels processos d'intercanvi de substàncies i defensa front als anticossos, bacteriòfags i fagocitosi. Únicament apareix en alguns tipus de bacteris.
La paret cel·lular bacteriana és rígida i presenta una estructura i composició diferent a la paret de les cèl·lules eucariotes vegetats. El component fonamental és el peptidoglicà, que es pot considerar com una molècula gegant en forma de xarxa Manté la forma del bacteri i permet el pas d'aigua i ions al seu través. Una imatge amb la sua estructura la teniu ací
La membrana plasmàtica presenta una estructura semblant a la de les cèl·lules eucariotes, tot i que varien algunes de les molècules presents. Les funcions bàsiques també són les mateixes. Una peculiaritat de la cèl·lula bacteriana és l'existència d'un conjunt de replecs membranosos interns que reben el nom de mesosomes. Els mesosomes incrementen la superfície membranal, s'uneixen al cromosoma bacterià i posseixen una gran quantitat d'enzims que regulen la replicació del cromosoma bacterià i processos metabòlics com la respiració cel·lular, fotosíntesi o fixació del N2
Els y funcionen de la mateixa forma que en les cèl·lules eucariotes, però sempre estan lliures pel citoplama i tenen lleugeres diferències de composició i grandària.
Les inclusions són substàncies que la cèl·lula sintetitza en èpoques d'abundància, o bé són residus del seu metabolisme.
Els flagells són estructures de locomoció formades per fibres de natura proteica, encara que la seua estructura és molt diferent a ls de les cèl·lules eucariòtiquesFisiologia dels bacteris.
Funcions de nutrició
Els bacteris són cèl·lules molt heterogènies,
ja que les diferents espècies poden dur a terme tots els
tipus de metabolisme possibles. Alguns poden utilitzar-ne, facultativament,
més d'un tipus en funció de les característiques
ambientals.
Els bacteris poden ser autòtrofs (fotosintètics
o quimiosintètics) , heteròtrofs (paràsits,
simbiòtics, sapròfits), aeròbics o anaeròbics
(estrictes o facultatius). aquests conceptes seràn estudiats més endavant.
Funcions de reproducció
La reproducció bacteriana es realitza mitjançant
bipartició (no mitosi), prèviament hi ha una duplicació
del materils genètic. Els bacteris també presenten
altres mecanismes, definits com a parasexuals en què intercanvien
material genètic (seràn estudiats més endavant)
La cèl·lula
eucariota està present en els altre regnes dels éssers
vius, protistes (algues i protozous),metazous ( animals), metàfits
(vegetals) i fongs.
La cèl·lula eucariota es caracteritza perquè
el citoplama manifesta un grau elevat de complexitat i organització:
el nucli, separat del citoplama per una membrana nuclear,
conté 2 o més molècules d'ADN bicatenari
lineal associat a histones, cosa que li permet empaquetar-se millor.
El citoplama conté nombrosos orgànuls i sistemes
de membranes com els mitocondris, cloroplasts, reticle endoplasmàtic,
etc, que estableixen tota una xarxa de compartiments cel·lulars
interrelacionats que es reparteixen les funcions metabòliques
(compartimentació cel·lular); això
permet més especialització i més eficàcia
en la realització de les seues funcions (hi poden tindre
lloc alhora reaccions contraposades en diferents estructures tancades
de la cèl·lula).
Un altre tipus de diferències entre ambdós tipus
de cèl·lules ve donat per l'estructura i funcionament
del seu material genètic. Les cèl·lules eucariotes
presenten una gran quantitat d'ADN, molt més del que necessiten
per a la codificació de les seues proteïnes: una cèl·lula
humana conté mil vegades més ADN que un bacteri,
però sembla ser que únicament una xicoteta quantitat
d'aquest (potser un 1% en les cèl·lules humanes)
conté la codificació de les proteïnes que són
produïdes (gens fragmentats: exons i introns). A més
a més, les cèl·lules eucariotes presents
als organismes pluricel·lulars presenten una gran quantitat
de gens que poden ser inactivats (diferenciació cel·lular)
| PROCARIOTES | EUCARIOTES | |
| Organismes | Bacteris, cianofícies, micoplasmes | Protistes, fongs, plantes i animals |
| Grandària | 1-10 mm de llarg | 10-100 mm de llarg |
| Metabolisme | Aeròbic o anaeròbic | Aeròbic (anaeròbic facultatiu) |
| Orgànuls | Pocs | Nucli, cloroplast, mitocondri, etc |
| ADN | Circular en el citoplama; no combinat amb histones; sense separació de la resta del citoplasma | ADN linial, molt llarg i que conté gens fragmentats (exons i introns); associat a histones; organitzat en cromosomes i envoltat d'una membrana nuclear |
| ARN i proteïnes | ARN i proteïnes sintetitzades al mateix compartiment | ARN sintetitzat i transformat al nucli; proteïnes sintetitzades al citoplasma |
| Citoplasma | Compartimentació citoplasmàtica escassa (mesosomes). | Sense citoesquelet, corrents citoplasmàtics ni exocitosis Compartimentació citoplasmàtica desenvo-lupada (orgànuls membranosos). Citosquelet format per filament proteics; corrents citoplasmàtics; endocitosi i exocitosi |
| Divisió cel·lular | Per bipartició o per espores | Per mitosi (o meiosi) |
| Organització cel·lular | Principalment unicel·lular | Principalment pluricel·lular amb diferenciació de les cèl·lules |
Altra taula comparativa, un poc més complexa, ja que inclou aspectes del funcionament del material genètic, la teniu ací
A continuació podem comparar l'estructura d'una cèl·lula procariota (bacteri) i de dues cèl·lules eucariotes ("animal" i "vegetal"). Noteu qye la cèl·lula procariota ha estat ampliada
 Cèl·lula procariota
Cèl·lula procariota
 Cèl·lula eucariota animal
Cèl·lula eucariota animal
 Cèl·lula eucariota vegetal
Cèl·lula eucariota vegetal
Altres esquemes de la cèl·lula eucariota animal i la la cèl·lula eucariota vegetal
Hom creu que tots els organismes
que viuen a hores d'ara sobre la Terra deriven d'una única
cèl·lula primitiva nascuda fa ara uns 3500
milions d'anys com a resultat d'un lent procés evolutiu
de les molècules orgàniques. Una fita important
al llarg d'aquest lent camí evolutiu va tenir lloc ara
fa 1.500 milions d'anys quan es va produir la transició
des de les petites cèl·lules amb estructura interna
relativament senzilla - procariotes-, fins les cèl·lules
eucariotes, més grans i complexes, tal com les trobem ara
als animals i les plantes.(veure apartat final de la teoria endosimbiòtica)
Les cèl·lules procariòtiques són simples
estructuralment però diverses es del punt de vista bioquímic:
hi ha espècies de bacteris que poden utilitzar com nutrients
qualsevol tipus de molècula orgànica - hidrats de
carboni, aminoàcids, greixos, etc- (heteròtrofs).
altres són capaços d'aprofitar com a font de matèria
el CO i el N2 de l'atmosfera (autòtrofs fotosintètics
i quimiosintètics)
Els bacteris es poden dividir ràpidament per fisió
binària: sota condicions òptimes una cèl·lula
procariota es pot dividir cada 20 minuts i per tant originar 4.1010 cèl·lules (xifra semblant a la població
humana mundial) en menys de dotze hores. Aquesta capacitat de
reproduir-se amb rapidesa permet que les poblacions de bacteris
s'adapten ràpidament als canvis ambientals mitjançant
mutacions espontànies i selecció natural.
Les primeres cèl·lules que van poblar la Terra van
ser problablement heteròtrofs que obtenien l'energia per
fermentació de les molècules orgàniques
presents a l'oceà primitiu (sopa primitiva) capaç
d'alimentar aquestes estructures dotades d'un metabolisme rudimentari.
D'aquesta manera els components moleculars de la "sopa primitiva",
no només van aportar els elements per a formar les cèl·lules,
sinó que també van subministrar l'energia necessària
per al seu manteniment i la seua autoperpetuació. Aquest
aliment molecular però, començà a escassejar
a mesura que anava creixent la població de cèl·lules
heteròtrofes. Sota aquestes condicions, alguns organismes
foren capaços d'utilitzar els àtoms de carboni i
nitrogen (en forma de CO2 i el N2) de l'atmosfera, la qual cosa els va atorgar
un clar avantatge evolutiu. CO2 i N2 són molt abundants, però
també són molt estables. Es necessita doncs una
gran quantitat d'energia, així com un gran nombre de reaccions
químiques complexes per transformar aquestes molècules
en molècules orgàniques.
En el cas del CO2 el mecanisme que va aparéixer fou
la fotosíntesi, procés en el qual s'aprofita
la radiació solar com a font il·limitada d'energia
per a la conversió de CO2 en compostos orgànics
com la glucosa. Aquest procés va modificar, a més
a més, la composició de l'atmosfera terrestre, a
causa de l'aparició d'una gran quantitat d'O2
(producte residual de la fotosíntesi) que s'hi acumulava
progressivament. A partir d'aquest oxigen es va formar l'ozò (O3), un derivat triatòmic que es forma per acció de la
radiació ultraviolada i les descàrregues elèctriques
sobre les molècules d'O2 i que té un important paper en la protecció dels éssers vius enfront de la radiació UV provinent de l'espai.
No obstant això, l'oxigen residual de la fotosíntesi
es va anar convertint a poc a poc en un verí mortal per
a les cèl·lules ja que els seus mecanismes de fermentació
anaeròbia eren destruïts per aquest gas. Molts d'aquests
organismes anaeròbics van desaparéixier; altres
van refugiar-se en zones innaccesibles a l'oxigen i han pervingut
fins ara. Un pas evolutiu revolucionari va ser la utilització
metabòlica de l'oxigen que fou utilitzat per les cèl·lules
per extraure més energia dels aliments mitjançant
un procés anomenat respiració aeròbia,
que desprén CO2 com a producte residual.
Quan es van acabar les reserves orgàniques de la sopa primitiva
aquests moderns heteròtrofs aerobis van "aprendre"
a obtenir nutrients de les molècules sintetitzades prèviament
pels fotosintetitzadors. Aquests dos tipus cel·lulars van
anar evolucionant conjuntament i han sabut reciclar els productes
residuals dels seu metabolisme: els heteròtrofs utilitzen
la pol·lució fotosintètica (oxigen) mentre
que els fotosintètics n'aprofiten el CO2 .
La vida és, doncs, un fenomen qui sap si inevitable en les condicions inicials i és, alhora , irrepetible, ja que quan va aparéixer a la Terra, va modificar l'atmosfera del planeta i va destruir les condicions ambientals que havien fet possible la seua pròpia aparició.
- 4500 ma Formació de la Terra
-3500 ma Primeres cèl·lules. Predomini dels bacteris procariòtics
-1500 ma Aparició de les cèl·lules eucariòtiques
-570 ma Explosió cambriana. Sorgiment dels éssers pluricel·lularsFig.13: Breu història de la vida en la Terra.
Un altre tipus d'estratègia
per sobreviure als canvis de composició atmosfèrica
fou la que tingueren diferents organismes anaeròbis que
s'associaren de forma íntima amb un tipus aeròbic
de cèl·lula, vivint ambdues en simbiosi. D'aquest
procés d'associació de diferents cèl·lules
es pensa que va sorgir la cèl·lula eucariota.
En la dècada dels '80, Lynn Margulis proposà la
teoria de l'endosimbiosi per explicar l'origen de mitocondris
i cloroplasts. segons aquesta teoria una gran cèl·lula
procariota englobaria altres cèl:lules procariotes menors,
fa ara -1500 ma.


Fig.14: Origen de
la cèl·lula eucariota per endosimbiosi
Els mitocondris mostren moltes semblances amb els organismes procariotes pel que fa a la seua grandària, forma, ADN i tipus de reproducció. Sembla probable que les cèl·lules eucariotes siguen les descendents d'organismes anaeròbics primitius que sobrevisqueren en un món qua havia canviat a ser ric en oxigen, mitjançant la incorporació de bacteris aeròbis. En lloc de digerir aquests bacteris, els van mantenir i alimentar ja que aquestes estructures tenien la capacitat per a consumir l'oxigen atmosfèric i produir-hi energia. Evidentment no podem demostrar d'una forma absoluta que allò va tindre lloc realment però alguns microorganismes actuals indiquen que aquesta seqüència evolutiva és factible: l'ameba Pelomixa palustris, no té mitocondris sinó que conté bacteris aeròbics en relació simbiòntica permanent. D'una manera semblant els actuals cloroplastos de les cèl·lules eucariòtiques fotosintètiques provindrien de l'associació simbiòntica de bacteris fotosintètics i cianobacteris.
Entrevista a Lynn Margulis en la revista MÈTODE de la Universitat de València

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}