El metabolismo energético matiene los abastecimientos de ATP y glucosa de dos formas:
1-cuando hay alimentos disponible, por medio de la formación de moléculas de almacenaje (glucógeno, grasas, proteínas)
2- por la recuperación de glucosa y ATP de este almacen cuando es requerido por el organismo.
La necesidad de glucosa o ATP puede constituir una demanda por cantidades masivas e inmediatas de energía o simplemente para mantener los niveles de energía y glucosa entre comidas.
INTEGRANDO LAS RUTAS METABOLICAS
Como ningún tejido puede sobrevivir metabolicamente sin la interacción con lo demás tejidos, este metabolismo energético es regulado por una extensa cooperación entre los diferentes órganos, pero siempre con esta función central: mantener los niveles adecuados de ATP y glucosa. Los cuatro tipos primordiales de tejidos, cada uno con su función metabólica especializada, son: hígado, músculos, adiposo y cerebro.
ATP
La hidrólisis de ATP es la fuente inmediata de energía para los procesos celulares. La principal fuente de ATP es la cadena de transporte electrónico (CTE), que ocurre en el mitocondrio, y que es alimentado por el ciclo de ácido cítrico (CAC), también conocido como ciclo de Krebs. Como la CTE requiere oxígeno, mucha de la producción de ATP está directamente relacionada con el suministro de oxígeno.
Como no tienen mitocondrio, los glóbulos rojos descansan totalmente en la glucólisis anaeróbica para obtener energía. Los músculos, por su parte, pueden ser forzados a descansar únicamente en la glucólisis anaeróbica cuando el ejercicio extenuante consume más oxígeno que el que puede se dispensado al músculo.
GLUCOSA
Los metabolitos que se producen de la degradación de glucosa son esenciales para la función del CAC. Para que esta ciclo continue funcionando es necesario mantener a un nivel razonable sus intermediarios. Hay que recordar que estos intermediarios se usan para la síntesis de otros compuestos ajenos al CAC, por ello hay que estar constantemente remplazándolos.
Piruvato se obtiene sólo de glucosa o de ciertos amino ácido. Las reacciones que convierten el piruvato en intermediarios del CAC se conocen como reacciones anapleróticas:
Piruvato ------> oxaloacetato
Reacción catalizada por carboxilasa de piruvato (carboxilasa dependiente de biotina)

Piruvato ------> malato
Racción catalizada por la enzima málica

El resultado de estas reacciones es la síntesis neta de todos los intermediarios del CAC, que son necesarios para remplazar a los intermediarios que son retirados del ciclo para la síntesis de otros compuestos.
Las células que no tienen mitocondrio (globulos rojos) tienen que usar glucosa para producir energía ya que no tienen ciclo de Krebs ni tampoco fosforilación oxidativa. Sin un constante suministro de glucosa estas células moririan. Otros tejidos, como el cerebro, dependen tambien del metabilosmo de glucosa para obtener la energia, sin embargo, el cerebro puede usar fuentes alternas de energía en caso de que la glucosa no esté disponible.
MOLECULAS DE ALMACENAMIENTO
Lo que pretende el metabolismo energético es el almacenamiento de glucosa y energía. Glucógeno, el polímero ramificado de glucosa que se acumula en el hígado, rińones y músculos, es un abastecedor, a corto plazo, de glucosa.
Por su gran cantidad y en términos de masa los mayores depósitos de glucógeno están en los músculos. Si embargo, los músculos almacenan glucógeno sólo para suplir sus propias necesidades. Como los músculos carecen de la enzima glucosa-6-fosfatasa, no pueden convertir glucógeno (o cualquier otro metabolito) en glucosa. El hígado y los rińones tiene esa actividad enzimática y comparten su depósitos de glucógeno para ayudar a mantener los niveles de glucosa en el cuerpo.
La energía se almacena primordialmente en forma de grasas. Las grasas se almacenan principalmente en el tejido adiposo, practicamente en cantidades ilimitadas. Se metabolizan por la oxidación Beta a acetil-CoA y luego, por el CAC, se queman completamente a CO2 para producir ATP.
Las grasas no se pueden usar para producir glucosa porque el acetil-CoA no puede convertirse directamente en los precursores de glucosa sin antes perder sus carbonos.
Las proteínas determinan los elementos estructurales y funcionales de las células, pero también se pueden usar para proveer energía. Las proteínas llevan a cabo un ciclo constante de síntesis y degradación. En tiempos de necesidad, la masa proteínica del cuerpo se puede usar para generar tanto glucosa como energía. Los amino ácidos derivados del desdoblamiento de las proteínas se pueden usar para producir energía o equivalentes de glucosa. Por ello, las proteínas son almacenes tanto de glucosa como de ATP.
ESTADOS METABOLICOS Y SUS SEŃALES
Consideremos estos tres estados metabólicos: alimentación, ayuno y estímulo o excitación, y tres principales seńales metabólicos: insulina, glucagón y epinefrina.
Alimentación: luego de cada comida los precursores de las moléculas de almacenamiento estan en cantidades abundantes; esto se conoce como estado postpandrial. Algo de este alimento se quema para suplir la energía inmediata, pero, por la acción de la insulina, la mayor cantidad se usa para el almacenamiento de ese alimento en forma de glucógeno, grasas y proteínas para uso posterior.
Ayuno: en este estado, parte de los depósitos de energía son reclamados por el sistema. Según disminuyen los niveles de glucosa, los niveles de insulina decaen y los de glucagón, la hormona que seńala bajos niveles de glucosa en la sangre, aumentan. Glucagón promueve la recuperación de energía de todas sus formas de almacenaje.
Estímulo: es el ímpetu de una inmediata necesidad de energía. Como respuesta a esta seńal de excitación, la médula adrenal secreta epinefrina a la circulación.
INSULINA
Luego de ingerir alimentos los niveles de glucosa aumentan y el páncreas secreta insulina. Como esta seńal implica altos niveles de glucosa en la sangre, la insulina promueve la entrada de glucosa en las células sensitivas a insulina. Esta hormona, secretada por las células beta del páncreas, se une a un receptor específico en la superficie de la célula para ejercer su efecto metabólico. A la vez que inhibe las rutas degradativas (degradación de glucógeno, grasas y proteínas), la insulina estimula el proceso de almacenaje: síntesis de glucógeno, grasas y proteínas.
GLUCAGON
Hormona que es la antítesis de la insulina, es producida por la células alfa del páncreas. Seńal que indica bajos niveles de glucosa sanguínea, estímula el desdoblamiento de glucagón, grasas y proteínas e inhibe su síntesis.
Glucagón aumenta la actividad de una específicas cinasas de proteínas celulares.Estas enzimas son las que usan ATP para fosforilar un residuo de serina, treonina y, ocasionalmente, tirosina de algunas proteínas específicas.
Cuando hay altos niveles de glucagón, unas proteínas específicas se fosforilizan. La fosforilación activa una enzimas específicas que tienen que activarse cuando las reservas de glucosa y energía son bajas y desactiva la enzimas responsables del alamcenamiento de energía.
Glucagón se une a un receptor en la superficie celular. Una vez ocupdo, el receptor, por la intercesión de una proteína de acoplamineto (una proteína G), activa la ciclasa de adenilato. Esta enzima toma ATP y forma AMP cíclico (cAMP) y fosfáto inorgánico, Pi. Al unirse el cAMP a la inactiva cinasa de proteína dependiente de cAMP se libera una subunidad inhibitoria y la enzima se activa. La activa cinasa de proteína comienza catalizar la fosforilación de otras proteínas, algunas de ellas cinasas también. El resultado neto es una gran amplificación de la seńal original (cinasas activando cinasas que a su vez activan otras cinasas) y un incremento en la fosforilación de proteínas celulares
La proteina G sirve com un cronómetro. Esta proteína atrapa GTP y, con el GTP unido, puede acoplar el receptor y activar la ciclasa de adenilato. La proteína G hidroliza lentamente el GTP a GDP y Pi; caundo esto ocurre, todo el complejo se desploma y la ciclasa de adenilato se inactiva.
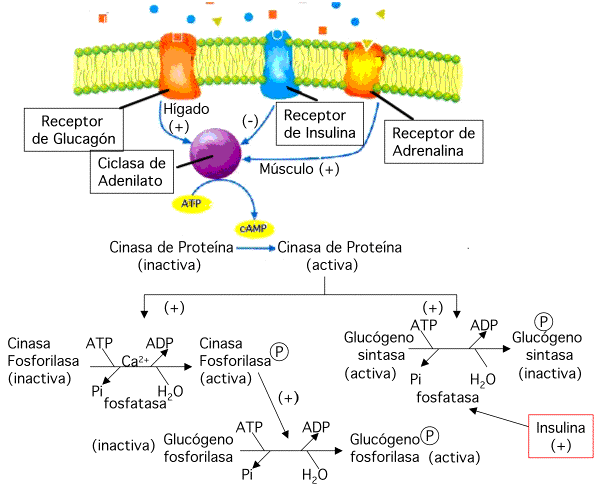
Cuando decienden los niveles de glucagón la enzima fosfodiesterasa de cAMP destruye el cAMP acumulado y unas específicas fosfatasas de proteína remueven el fosfáto de las fosfoproteínas. Usualmente, estas mismas fosfatasas son reguladas por la fosforilación. [Hay cinasas de fosfatasas (fosforilizan las fosfatasas) y fosfatasas de fosfatasa (desfosforilizan las fosfatasas]
En resúmen: aumento en los niveles de glucagón llevan a un incremento en la fosforilación de proteínas y disminución en los niveles de glucagón a un decenso en la fosforilación de proteínas.
EPINEFRINA
Hormona producida por la médula adrenal y sistema nervioso simpatético, impulsa a una rápida mobilización de energía y glucosa. Como el glucagón, epinefrina se une a un ceceptor celular específico y activa la ciclasa de adenilato. De aquí que su efecto sea similar al del glucagón: estímula el desdoblamiento de glucagón, grasas y proteínas e inhibe su síntesis.
SEŃALES SECUNDARIAS
Las seńales hormonales primarias sirven como seńales extracelulares que son interpretadas por un aparato de transducción de seńal que las convierte en seńales intracelulares o mensajeros secundarios que, a su vez, "avisan" a enzimas individuales dentro de la célula de lo que ocurre fuera de ésta. Estos mensajeros secundarios se pueden agrupar en cuatro categorías:
Seńales de alta energía = ATP, citrato, ácidos grasos, NADH, acetil-CoA
Seńales de baja enrgía = cAMP, AMP, ADP, Pi
Seńales de alta glucosa = Fructosa-2,6-biP, glucosa-6-P
Seńales de baja glucosa = cAMP
No todas estas moléculas afectan todas las enzimas y/o las rutas metabólicas, sino que, de tener algún efecto, será en la dirección indicada por el tipo de seńal. Por ejemplo, fructosa-2,6-biP es seńal de altos niveles de glucosa. Por ello, es de esperar que una alta concentración de fructosa-2,6-biP aumente el metabolismo de glucosa a través de glucólisis, aumente la síntesis de ácidos grasos, y aumente el almacenamiento de glucógeno y proteínas. Sin embargo todo lo que hace la fructosa-2,6-biP es aumentar la actividad de glucólisis (activando la fosfofructocinasa), y disminuir la gluconeogénesis (inhibiendo la fructosa-1,6-bifosfatasa), pero no afecta directamente la actividad de otras enzimas.
GENERALIDADES DEL METABOLISMO
1- LOS NIVELES DE ATP Y GLUCOSA DEBEN SER RAZONABLEMENTE CONSTANTES
2- SE REQUIRE GLUCOSA PARA METABOLIZAR LAS GRASAS Y PRODUCIR ENERGIA
Las grasas producen mucho ATP, pero éstas no se pueden metabolizar sin la presencia de carbohidrátos. Glucosa, o los intermediarios del CAC, no pueden sintetizarse a partir de acetil-CoA. La única forma de obtener intermediarios del CAC es de glucosa o de amino ácidos (proteínas). Como los intermediarios del CAC se usan para la generación de otros compuestos, están constantemenmte consumiéndose y tienen que ser contínuamente repuestos para mantener corriendo el ciclo. Si hay suficiente glucosa (o glucógeno), los intermediarios del CAC oxaloacetato y malato se pueden producir direactamente de piruvato (reacciones anapleróticas). Pero si no hay suficiente glucosa, los derivados del CAC deben obtenerse de la degradación de amino ácidos (proteínas). Por lo tanto, sin glucosa no hay catabolismo de grasas para producir energía.
3- GLUCOSA NO SE SINTETIZA PARTIENDO DE GRASAS
El producto final del metabolismo de grasas es acetil-CoA, y éste no se puede usar para generar glucosa porque para ello hace falta oxaloacetato, que sólo se puede producir a partir de acetil-CoA por el CAC con oxaloacetato.
Si no hay glucosa de la dieta ésta se obtiene de las proteínas y del glicerol de la degradación de triacilgliceroles.
4- SINTESIS Y DEGRADACION NO PUEDEN OCURRIR SIMULTANEAMENTE
5- BAJOS NIVELES DE ENERGIA ACTIVAN LA GLUCOLISIS Y LA LIPOLISIS
El CAC acoplado a la fosforilación oxidativa mitocondrial es la forma primaria de producir ATP. El acetil-CoA destinado al CAC se puede obtener del metabolismo de glucosa (glucólisis) o por la degradación de grasas (oxidación beta). Por lo tanto, bajos niveles de energía mobilizan los depósitos de glucógeno y lípidos.
6- BAJOS NIVELES DE GLUCOSA ACTIVAN LA GLUCONEOGENESIS Y LA DEGRADACION DE PROTEINAS
Para mantener los nivéles sanguíneos de glucosa hay que degradar glucógeno o sintetizar glucosa a partir de piruvato (gluconeogénesis). El glucógeno se almacena en el hígado y los músculos. La gluconeogénesis ocurre primordialmente en el hígado y los rińones. Si los niveles de glucosa sanguíneo son bajos, el hígado y los rińones suplen gucosa a la sangre por glucogenólisis y por gluconeogénesis. El glucógeno tiene diferentes funciones en diferentes tejidos. En hígado y rińones se degrada para suplir glucosa al resto del cuerpo o se puede usar para energía. En los músculos el glucógeno sólo se usa localmente para generar energía via glucólisis. Los músculos esquelatales no producen glucosa a partir de glucógeno porque carecen de la enzima glucosa-6- fosfatasa.
7- LA FOSFORILACION DE PROTEINAS COMO RESPUESTA AL AUMENTO EN LOS NIVELES DE cAMP ACTIVAN ENZIMAS QUE MANTIENEN LOS NIVELES DE GLUCOSA Y RECUPERAN ENERGIA E INACTIVAN LA ENZIMAS QUE ALMACENAN GLUCOSA, GRASAS Y PROTEINAS.
Bajos niveles de energía y glucosa están asociados al incremento en la actividad de cinasas de proteínas dependiantes de cAMP y a un aumento en la fosforilación de proteínas.
La fosforilación -desfosforilación en los residuos de serina o treonina es uno de los medios promordiales para controlar la actividad enzimática. La fosforilación de proteínas es una reacción dependiente de ATP y esta catalizada por numerosas cinasas:
Proteína-OH + ATP -----> Proteína-O-P + ADP
Esta modificación no es directamente reversible, pero el grupo fosfáto se puede remover de la proteína por la acción de una fosfatasa.
Proteína-O-P + ADP -----> Proteína-OH + Pi
La fosforilación activa algunas proteínas y desactiva otras. La actual fosforilación de una proteína regulada casi siempre está catalizada por una cinasa de proteína que es específica para sólo una o sólo unas pocas proteínas. Por otro lado, las mismas cinasas son comunmente reguladas por mecanismo de fosforilación-desfosforilación.
En el metabolismo energético lo que inicia todo el proceso de fosforilación es la cinasa de proteína dependiente de cAMP. Esta enzima es activada por un aumento en los niveles de cAMP, un mensajero secundario de baja energía y bajos niveles de glucosa. La activación de la cinasa de proteína dependiente de cAMP es la que lleva a un aumento en la fosforilación de proteínas.
Como regla general, las enzimas requeridas sólo durante condiciones de baja energía y baja glucosa se activan por fosforilación y las enzimas requeridas para el proceso contrario (condiciones de alta energía y alta glucosa) se desactivan por fosforilación.
Ejemplo : la fosforilación de fosforilasa de glucógeno activa la enzima, que es la responsable de degradar glucógeno a glucosa-1-P. La degradación de glucógeno en hígado y músculos es requerida bajo condiciones de baja glucosa o baja energía, condiciones asociadas con un aumento en la fosforilación de proteínas. Como contraste, la enzima responsable de la síntesis glucógeno y que debe estar inactiva bajo condiciones de bajos niveles de glucosa, la sintasa de glucógeno, es inactivada por fosforilación.
Ahora bien, la fosforilación para activar o inactivar enzimas tambien depende del tejido en el que esta ocurriendo la ruta metabólica. Por ejemplo, en el hígado cAMP activa gluconeogénesis pero en el músculo activa glucólisis.
Veamos el proceso desde el punto de vista de la fosfofructocinasa-2 (PFK-2), la enzima que cataliza la síntesis de fructosa-2,6-bifosfáto a partir de fructosa-6-fosfáto:
Fructosa-6-P --(PFK-2)--> Fructosa-2,6-biP.
La reacción inversa está catalizada por fructosa bifosfatasa-2 (FBPasa-2):
Fructosa-2,6-biP --(FBPasa-2)--> Fructosa-6-biP.
La actividad enzimática de la PFK-2 y la FBPasa-2 está localizada en diferentes dominios de una misma proteína (proteína bifuncional).
Alta conecentración de Fructosa-2,6-biP estimula glucólisis por la activación alostérica de fosfofructocinasa-1(PFK-1) que es la enzima que cataliza la reacción glucolítica:
Fructosa-6-P --(PFK-1)--> Fructosa-1,6-biP.
La reacción inversa está catalizada por fructosabifosfatasa-1 (FBPasa-1):
Fructosa-1,6-biP --(FBPasa-1)--> Fructosa-6-P.
Baja concentración de Fructosa-2,6-biP estimula gluconeogénesis por la activación alostérica de fructosabifosfatasa-1 (FBPasa-1) que es la enzima que cataliza la reacción gluconeogénica.
En el hígado, si los niveles de cAMP son altos entonces ocurre la fosforilación de PFK-2 para desactivarla y la desfosforilación de FBPasa-2, para activarla. El resultado neto es la diminución de los niveles de fructosa-2,6-biP, lo que hace que la glucólisis se desactive y se dé la gluconeogénesis.

En el músculo, si los niveles de cAMP son altos entonces ocurre la fosforilación de PFK-2 para activarla y la desfosforilación de FBPasa-2, para desactivarla. El resultado neto es un aumento de los niveles de fructosa-2,6-biP, lo que hace que la glucólisis se active (En los músculos no se dá la gluconeogénesis).

MOVIMIENTOS METABOLICOS DE GLUCOGENO

GRASAS
Proveen una forma de almacenamiento de energía (ATP) a largo plazo.
Hígado = síntesis de grasas para exportar a otros tejidos.
Músculo = en receso prefiere ácidos grasos como fuente de energía. Glucosa es la fuente de energía inmediata.
Tejido Adiposo = principal depósito de grasas. Una lipasa sensitiva a hormona mobiliza los triacilgliceroles (triglicéridos) y los hidroliza a ácidos grasos libres y glicerol.
Cerebro = no usa las grasas como fuente energía, pero en caso de inanición usa los cuerpos cetónicos como combustible.
MOVIMIENTOS METABOLICOS DE LAS GRASAS

METABOLISMO DE GRASAS

PROTEINAS
Reserva a largo plazo de glucosa y energía. Tambien son importantes componentes estructurales de las células. Uso prolongado de proteínas para generar energía y glucosa merman la masa muscular.
Hígado = depósito menor de proteínas pero ayuda al músculo a metabolizar las proteínas.
Músculo = principal depósito de proteínas para las necesidades metabólicas.
Tejido Adiposo = no es un depósito significativo de proteínas.
Cerebro = no es un depósito significativo de proteínas.
MOVIMIENTO METABOLICO DE PROTEINAS

METABOLISMO DE PROTEINAS

Las proteínas no son únicamente un embalse para el almacenamiento de energía y glucosa, sino que son componentes estructurales esenciales y funcionales de las células. No hay una degradación rampante y no específica de proteínas: el proceso es selectivo. Algunas proteínas se degradan rápidamente aún bajo condiciones normales. Este tipo de sistema de síntesis-degradación se usa como un mecanismo para controlar las rutas metabólicas que usan las proteínas como enzimas. El sistema cíclico de síntesis-degradación no provee de una cantidad significativa de amino ácidos para atender las demandas metabólicas; las proteínas se degradan para proveer para estas necesidades sólo como último recurso.
El cuerpo tiene que generar una fuente de equivalentes de glucosa para su uso. Los equivalentes de glucosa son esenciales. Las grasas no proveen equivalentes de glucosa. Cuando hay una ingestión limitada de glucosa (ayuno) o una capacidad limitada para utilizarla (diabetes), las proteínas se degradan para proveer ese esencial abastecimiento de glucosa.
Cada amino ácido tiene su propia ruta degradativa. Algunos se degradan a piruvato o a algún intermediario del CAC, y se conocen como amino ácidos glucogénicos. De éstos es que se obtiene glucosa cuando no está disponible.
Dos de los amino ácidos (Leucina y Licina) son cetogénicos: se degradan sólo a acetil-CoA. Estos no se usan para la generación de glucosa. Algunos amino ácidos son tanto glucogénicos como cetogénicos.
Las proteínas se pueden degradar a amino ácidos no-esenciales los cuales se degradan para suplir amino ácidos esenciales para la síntesis de proteínas. Esto ocurre en casos de deficiencia de amino ácidos esenciales en la dieta. Este proceso lleva a una gran degradación de proteínas y a una sobreabundancia de amino ácidos no-esenciales y sus metabolitos (balance negativo de nitrógeno). El exceso de aminacidos debe degradarse; el nitrógeno liberado se elimina como amoniaco y urea.
La localización primaria de depósitos de proteínas metabólicamente útiles son los músculos. Mientras haya suficiente glucosa las proteínas no son degradadas. No obstante, normalmente las proteínas están en un constante proceso balanceado de síntesis y degradación (balance de nitrógeno = 0).
CICLOS INTERORGANOS
Ciclo de Cori = músculos usan glucosa proveniente del hígado para hacer lactato (condiciones anaeróbicas). El hígado usa el lactato de los músculos para hacer glucosa.
Ciclo de Alanina = hígado toma esqueletos de carbono y deshechos de nitrógeno del músculo (alanina), dispone del nitrógeno (ciclo de urea) y recicla el carbono en glucosa.
Cuerpos Cetónicos = acetoacetato y B-hidroxibutirato. Se forman en el hígado (de la oxidación beta) para liberar CoA. Se metabolizan en otros tejidos como fuente de energía. El cerebro, bajo condiciones de inanición o diabetes, induce la enzimas a uasarlas como fuentes de energía.