La función primordial de metabolismo es el reciclaje de ATP, aunque la formación de éste no está directamente acoplado a las reacciones metabólicas: más bien se produce por la oxidación de NADH y FADH2, que se producen durante las reacciones catabólicas.
El anabolismo usa la energía almacenada en el ATP para realizar trabajo. y al catabolismo regenera, indirectamente, el ATP.
ETAPAS DEL METABOLISMO

Catabolismo:
Etapa I - degradación de macromoléculas (proteínas, grasas, (traicilgliceroles), polisacáridos) en sus respectivos componentes: aminoácidos, ácidos grasos, glicerol, y monosacáridos (azúcares).
Etapa II - Los productos de la primera etapa se convierten en intermediarios metabólicos y en un metabolito común: AcetilCoA. En esta etapa se libera algo de enrgía en forma de NADH y ATP.
Etapa III - AcetilCoA entra en el ciclo de ácido cítrico (Krebs) donde se oxida a CO2, que es el producto final del metabolismo aeoróbico de carbono. Aquí se producen los cofactores reducidos reactivos NADH y FADH2 que, eventualmente, ceden sus electrones. Estos electrones son transportados hacia oxígeno (O2) por la cadena respiratoria. La energía liberada por el transporte de electrones está acoplada a la síntesis de ATP el cual se genera a partir de ADP y Pi. El proceso se conoce como fosforilación oxidativa.
Las tres rutas principales convergen en una vía común, típico del catabolismo. En el anabolismo las rutas principales son divergentes: comienzan con diferentes contratos o intermediarios. Muchas de estas vías necesitan energía en forma de ATP o NADPH.
CATEGORIAS DE LAS REACCIONES BIOQUIMICAS
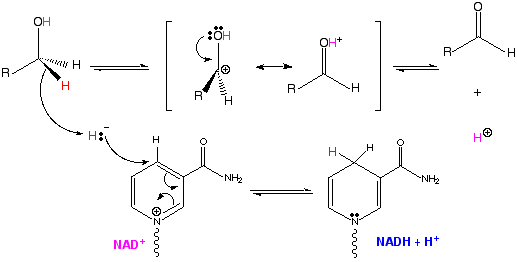
(1) OXIDACION-REDUCCION = catalizadas por Oxidorreductasas (deshidrogenasas) - Catalizan la transferencia de electrones.
Son las más comunes; se transfieren electrones de una molécula o átomo a otra molécula o átomo, por lo que tiene que haber dos reactantes: uno para donar electrones (agente reductor) y otro para aceptarlos (agente oxidante):
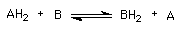
AH2, el agente reductor (pierde electrones), se oxida.
B, el agente oxidante (gana electrones), se reduce.
En muchas reacciones bioquímicas los átomos de carbono en los sustratos son usualmente el foco de los procesos de oxidación reducción:
-Oxidación: el carbono forma enlaces con carbono o nitrógeno, o cuando pierde hidrógeno (la densidad electrónica del carbono disminuye).
-Reducción: el carbono aumentan los enlaces con hidrógeno o disminuye los enlaces con oxígeno o nitrógeno (la densidad electrónica del carbono aumenta).
La oxidación-reducción en estos carbonos se puede indicar, también, por el cambio en hibridación o geometría molecular: los de mayor estado de oxidación son planos (sp2) o lineales (sp).
Carbonos con hibridición sp3 están en el más bajo nivel de oxidación (más reducidos).
La oxidación supone una reducción y viceversa. En bioquímica, muchas reacciones redox están acopladas a los pares de coenzimas redox: NAD+/NADH; NADP+/NADPH; y FAD/FADH2. Por ejemplo:
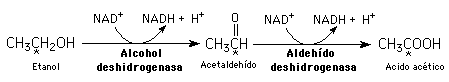
Etanol y acetaldehído son donantes de electrones (agentes reductores); NAD+ es el agente oxidante porque acepta los electrones.
Metales ionico, especialmente las parejas de cobre (Cu+/Cu2+) y hierro (Fe2+/Fe3+), muchas veces asociados con enzimas y otras proteínas, también funcionan como agentes oxidantes y reductores:
Cu+ <===>Cu2+ + e-
(2) TRANSFERENCIA DE GRUPOS= catalizadas por Transferasas - Catalizan la transferencia de grupos funcionales de una molécula a otra (intermolecular) o en la misma molécula (intramolecular). Ejemplos de grupos transferibles: fosforilo (-PO32-), amino (-NH2); carboxilo (-COOH); carbonilo (C=O); metilo (-CH3); y acilo (R-C=O).
Por ejemplo, la transferencia de un grupo fosforilo de ATP a glucosa está catalizada por una fosfotransferasa, mejor conocida como hexocinasa:

Otro ejemplo es la transferencia del grupo acilo a coenzima A (CoASH) formando un enlace tioéster entre el grupo acilo y el -SH de CoASH:
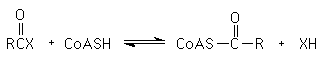
(3) RUPTURA HIDROLITICA (HIDROLISIS) = catalizadas por Hidrolasas - Catalizan la ruptura de ciertos enlaces por hidrólisis (transferencia de los grupos funcionales de agua).
En estas reacciones se utiliza agua para la hidrólisis de:
(a) Esteres - triacilgliceroles a ácidos grasos:

(b) Amidas - enlaces péptidos, por medio de peptidasas:
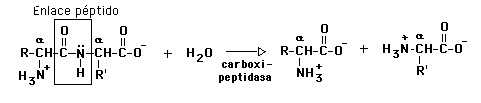
(c) Glucósidos - oligosacáridos a monosacáridos:

(4) RUPTURA NO-HIDROLITICA = catalizada por Liasas - Catalizan la formación de dobles enlaces por eliminación de grupos o la adición de grupos a un doble enlace (saturación), y también la escisión de una molécula por procesos no hidrolíticos.
La reacción más frecuente es la de la ruptura del enlace C-C por liasas:

La reacción de remoción de grupos para formar dobles enlaces (reacción directa, en la figura) o la adición de grupos a dobles enlaces (reacción inversa, en la figura) se ilustra a continuación:

(5) ISOMERIZACION Y REARREGLO - catalizada por Isomerasas - Catalizan la transferencia de grupos dentro de una molécula (rearreglos intramoleculares) para dar formas isoméricas.
Hay dos tipos; en cada uno se transforma una molécula del sustrato en su isómero:
1- desplazamiento intramolecular de átomos de hidrógeno (H.): cambia la posición de dobles enlaces (C=C, C=O, etc.)

2- rearreglos intramoleculares de grupos funcionales (poco frecuentes en el metabolismo).

(6) FORMACION DE ENLACES USANDO ENERGIA (ATP) - catalizada por Ligasas o Sintetasas - Catalizan la formación de enlaces C-C, C-S, C-O, y C-N por condensación acoplada con la ruptura (hidrólisis) de ATP, que suple la energía necesaria.
Para los enlaces C-C se usan carbaniones estabilizados con grupos carbonilos de cetonas, ésteres o CO2:

El carbanión se produce a partir de un grupo acetilo y se estabiliza por resonancia:
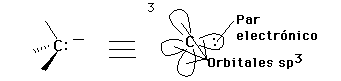
Por ejemplo:

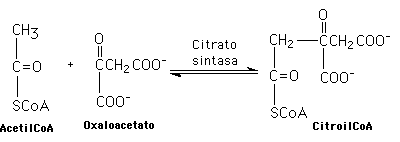
Muchas reacciones metabólicas surgen de la combinación de dos o más de la seis ilustradas en un sólo paso metabólico. Un ejemplo es la descaboxilación de isocitrato (descarboxilación oxidativa):

BIOENERGETICA
La energía puede visualizarse como la capacidad de generar o experimentar un cambio. El metabolismo es un proceso de transformación de energía donde el catabolismo proporciona la energia requerida para el anabolismo.
La forma de intercambio energético es la molécula de trifosfáto de adenosina (ATP), que es el transportador universal de energía en los organismos vivos.
El catabolismo es un proceso en el que se libera energía: es espontáneo. El anabolismo, por su parte, requiere energía.
Fundamentalmente sólo hay dos razones por la que ocurre una reacción química:
(1) la tendencia a lograr el mínimo de energía
(2) la tendencia de lograr el máximo de desorden.
El que una reacción ocurra a una temperatura dada dependerá del balance entre estas dos tendencias.
Estas tendencias lo que implican es que la espontaneidad de una reacción química se puede determinar relacionando las propiedades termodinámicas de entalpia
( H) y entropía (S).
H) y entropía (S).
El cambio en entalpía nos dá una medida del calor ganado a presión constante; está relacionado a la energía requerida para romper enlaces químicos.
La entropía, por su parte, es una medida de la menera que un sistema se rearregla; está relacionada con el grado de dispersión de la materia y la energía.
Las reacciones químicas se pueden entender en términos de estos dos efectos: la tendencia a formar los enlaces más fuertes posible, y la tendencia a dispersarse (mayor desorden). La dirección y la extensión de la reacción es la resultante de estas dos tendencias.
De acuerdo a la termodinámica, la dirección preferencial para una reacción está determinada por el compromiso entre las tendencias hacia enlaces más fuertes y mayor desorden.
Se dice, entonces, que la diferencia entre H y TS equivale a una cierta cantidad de energía útil para hacer trabajo, la que se denomina como energía libre y se simboliza con la letra G. Esta propiedad del sistema se conoce comunmente como la energía libre de Gibbs.
En realidad, lo que se determina es el cambio en esta energía libre (G), aplicando la siguiente ecuación:
G = H - TS Un decenso en la energía libre implica la cantidad de energía que está disponible (libre) para hacer trabajo. Es decir, es la energía que "sobra" luego de concluído todo el procreso.
Como el decenso en energía libre es trabajo útil que puede obtenerse de un proceso a presión y temperatuta constante, se puede usar como la medida de la espontaneidad del proceso. (medida de la tendencia del proceso a proceder espontáneamente, cuando el proceso se lleva acabo a presión y temperatura constante).
Si se considera un cambio en un sistema y se calcula un decenso en la energía libre para ese cambio, se puede esperar que ese cambio sea el producto de un proceso epontáneo.
Por lo general, los valores de G se intrepretan como sigue:
G negativo (<0): reacción es espóntánea en la dirección descrita (exergónica). Reactantes tienen más energía que los productos, se libera energía útil para hacer trabajo.
G positivo: (>0): la reacción es epontánea en la dirección contraria a la descrita (endergónica). Reactantes tienen menos energía que los productos, se requiere energía para que la reacción proceda según se indica (directa).
G = 0 : el sistema está en equilibrio.
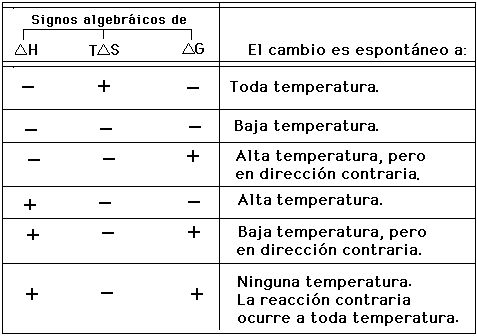
La predicción de la espontaneidad de un sistema se determina por el decenso en la energía libre de Gibbs, es decir, cuando el valor de G es negativo. Para hacer la predicción es necesario conocer los signos algebráicos de H y TS, dado que:
G=H - TS
La siguiente tabla sirve para estos propósitos:
COMPUESTOS FOSFATADOS DE ALTA ENERGIA
Por sus enlaces fosfoanhídros ATP es considerado como uno de los compuestos fosfatados de alta energía:
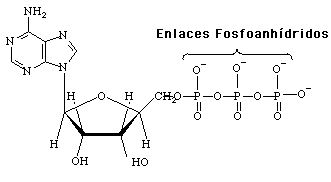
Los compuestos fosfatados de alta energía tienen una muy alta energia libre negativa a causa de su hidrólisis; para ATP se liberan 31 kJ/mol debido a la ruptura de los enlaces fosfoanhídros:
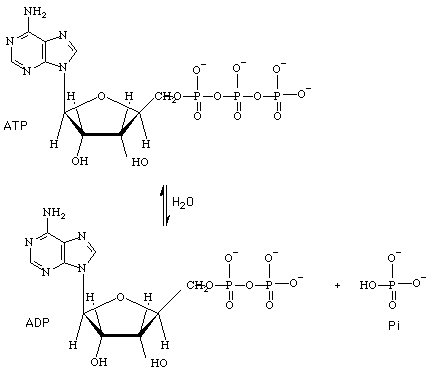
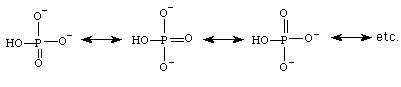
ATP tiene una alta potencialidad para transformarse en ADP + Pi. Como éstos últimos están en un más bajo nivel energético que ATP, se libera energía. ADP y Pi son mas estable que ATP por su resonancia (tienen más formas resonantes que ATP y agua) y menor repulsión de cargas negativa. Precisamente porque en ATP hay mayor repulsión de carga negativa es que aumenta la posibilidad de que se rompan los enlaces fosfoanhídros.

ATP y ADP son moléculas reactivas o ricas en energía, pero no son las únicas; por ejemplo, fosfoenolpiruvato (PEP) es también un compuesto fosfatádo de alta energía: su hidrólisis tiene un G° de -61.9 kJ/mol:

Mientras más alto el G° mayor el potencial de transferencia del grupo fosforilo (Pi). esto es una medida de la tendencia relativa de transferir un grupo fosforilo a una molécula aceptadora. A continuación se ofrece una lista de algunos compuestos fosfatádos de alta energía (el G° de hidrólisis, en kJ/mol, se ilustra en el paréntesis):
Fosfoenolpiruvato (-61.9)
1,3-bifosfoglicerato (-49.3)
Fosfocreatina (-43.0)
ATP (-30.5)
ADP (-30.5)
Glucosa-1-fosfáto (-20.9)
Glucosa-6-fosfáto (-13.8)
Reacciones termodinamicamente desfavorables pueden hacerse favorables si se acoplan con reacciones con un alto valor negativo de G°. En organismos vivos, la hidrolisis de compuestos fosfatádos se usa para esos propósitos, siendo ATP uno de los más empleados, por dos razones primordiales:
1- tiene una alta capacidad de transferencia de Pi
2- puede actuar como intermediario común para enlazar energéticamente dos reacciones.
Ejemplo A:
PEP ⇔ Piruvato + Pi ----
ADP + Pi ⇔ ATP ----G° = -61.9 kJ/mol (termodinamicamente favorable).G° = +30.5 kJ/mol (termodinamicamente desfavorable).
Si se acoplan ambas, la reacción neta será:
PEP + ADP ⇔ Piruvato + ATP ----G° = -31.4 kJ/mol (termodinamicamente favorable).
Ejemplo B:
ATP ⇔ ADP + Pi ----
Glucosa + Pi ⇔ Glucosa-6 fosfáto ----G° = -30.5 kJ/mol (termodinamicamente favorable).G° = +14.0 kJ/mol (termodinamicamente desfavorable).
Si se acoplan ambas, la reacción neta será:
ATP + Glucosa ⇔ Glucosa-6 fosfáto + ADP ----G° = -16.5 kJ/mol (termodinamicamente favorable).
Ejemplo C:
Las siguiente reacción no se produce directamente porque no hay enzima que la catalice:
PEP + Glucosa → Glucosa-6-fosfáto + Piruvato
Sin embargo, acoplando las siguientes se puede obtener la anterior:
PEP + ADP ⇔ Piruvato + ATP ----
ATP + Glucosa ⇔ Glucosa-6 fosfáto + ADP ----G° = -31.4 kJ/mol (termodinamicamente favorable).G° = -16.5 kJ/mol (termodinamicamente favorable).
PEP + Glucosa → Glucosa-6-fosfáto + Piruvato ----G° = -47.9 kJ/mol (termodinamicamente favorable).
Si no se produce ATP no habrá manera de transferir energía para las reacciones biosintéticas u otros procesos que requieran energía. De aquí que ATP se considere como el reactivo universal para la transferencia de energía metabólica.