Mirogrex terraesanctae biomass
changes, their causes and ecological impact
Sections:
·
History
·
Causal factors
·
Ecological impact
History of biomass changes
Surface area of Lake Kinneret
- 167.8 sq. km
A- Drainage of Lake
Hula, 1953 – 1957
B - Settling ponds begin operation in Kinneret catchment, 1982
C - Level of L. Kinneret lowered to nearly -213 m,
1990 and 1991
D - Epidemic of ectoparasites on lavnun, 1998
The earliest available data on Kinneret sardine catches and catch composition are those
recorded by British Mandate authorities in the 1930's when annual yield was
~100 tonnes. In the war years of the 1940's the catch
rose to ~250 T/a. This was a throw-net fishery and catches contained a high
proportion of small specimens (Ricardo-Bertram,
1947; Reich, 1978).
After the establishment of the State of
Israel in 1948, purse seining with lights (known as 'sekalawa')
was introduced, and a 'minimum legal size' of 12 cm total length was enforced.
The catch shot up, reaching ~700 T in 1952, but in spite of increased fishing
effort, it fell and leveled off at ~550 T/a in the next 7 years (Reich, 1978),
implying that the fishery was taking maximum sustainable yields in the 1950’s.
Thus, according to Beverton & Holt’s 1966 population model, total
stock in the exploited phase was ~1700 T, or ~2000 T
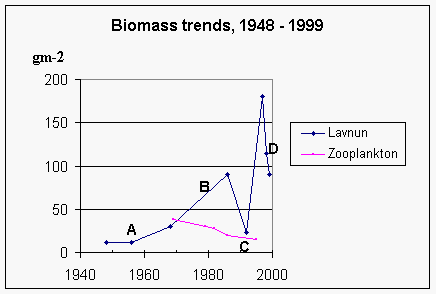
with the smaller specimens included (Landau, 1991.and Figure entitled 'Biomass trends' given above).
As noted by Reich (1978) there was an
unexpected rise in the sardine catch to ~800 T/a in the early 1960’s. From the growth hypothesis
it can be deduced that this abundance derived from broods hatched between 1953
and 1956, when Lake
Hula in the northern
watershed of L. Kinneret was being drained.
With improved fishing strategy the catch was
brought up to 1000 T/a by 1968. However, due to marketing difficulties that
began in the 1970’s (Reich, 1978) the sardine fishery took only ~1000 T/a
between 1968 and 1986. But catch-per-unit-effort data of Israel Dept. of
Fisheries show a fourfold increase in standing stock abundance in this period.
At its peak in the mid-1980’s, the stock amounted to
15000 – 20000 tonnes, including the unexploited
portion. This estimate is based on various sources: the 1950’s estimate of 2000
T, catch and effort analysis, and mortality estimates from age composition data
(Landau, 1991).
In the1986/87 fishing season, the commercial
stock reached an unprecedented biomass,
indicated by a mean catch-per-unit-effort of 5.3 T, twice the average C/E of
the previous 5 years (Landau, 1996). At the same time, the scarcity of young
specimens in the catches indicated imminent collapse due to recruitment
failure.
Following a decline in C/E (Landau, 1991;
1996) the commercial catch dropped to 300 T in 1994. The exploited portion of
the stock may have been as low as 1000 T (a more accurate estimate cannot be
given because data on effort are not available).
In December 1993, the ‘dilul’
or ‘thinning’ project got underway,
using a mesh-size which retained specimens over 8 cm total length.
Between 1993 and 1997 biomass increased along with fish size, bringing total
stock up to ~30,000 T (Landau, 1998
and 'dilul
') The
main components of this super-abundant stock were two broods, those of 1990/91
and 1991/92, hatched when the level of L. Kinneret
approached -213 m for the first time in1990 and the second time in December
1991 (Landau, 1997 , Mirogrex
growth).
In 1998 the sardine stock became heavily
infested by the ectoparasite Lernea
cyprinacea (Copepoda).
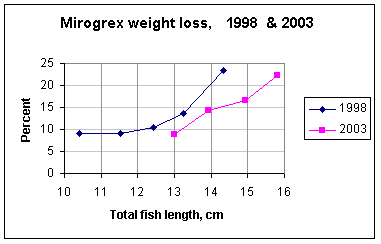
Among the larger specimens (13 – 14 cm TL) over 50% had lesions or attached
parasites, and weight loss averaged 18%; there was less infestation and weight
loss among the specimens 10.5 – 12.5 cm TL. Calculation of
weight loss is based on data for former years (Landau, 1991).

Update. 2003
Two samples of commercial catch totalling 115 specimens were taken in March 2003.
Their condition was slightly better than that of 1998 specimens in the same
size range, but mean weight was still 15% lower than that of Mirogrex of earlier years (diagram below). Signs of
parasitic infestation were seen in 26 of the 75 specimens inspected.
Ostrovsky (report T8/2003, Israel Oceanographic & Limnological Research) presented data showing a low
condition factor for all size categories in 1998, 1999 and 2000, with especially
low values for larger fish; 2001 values conform to the norms of the early
1990's. Neither this report nor any other as far as I know, mentions the
epidemic of parasites, or attempts to explain observed weight losses.
Larger specimens were under-represented in
the 1998 size distribution, denoting mortality due to infestation (Landau,
1998; Figure entitled 'Size and age distribution of lavnun
in the 'dilul' fishery'). Stock decline is apparent
from the 1999 catch and effort data of the Kinneret
Authority given in the 'dilul
'
Webpage. Later C/E data is not available.
In fishponds under natural
conditions, epidemics of parasites occur when population density reaches 200
kg/ dunam (1000 sq. m.)(personal
communication, Dr. Bejarano, Fish Disease Lab., Nir David). For an area the size of L. Kinneret,
167.8 sq. km., 200 kg/dunam amounts to 33,000 tonnes. Thus the parasite epidemic confirms the stock
estimate of ~30,000 T, and demonstrates that this is the carrying capacity of
the lake.
Factors of biomass change
The survival of Mirogrex
larvae depends upon their ability to find patches of suitable food when the
yolk-sac is depleted. As in many other fish species, survival at this critical
stage is a major determinant of population size. Studies in the 1980's
demonstrated the importance of small organisms, especially the rotifers Synchaeta spp. (Landau et
al, 1988 and others). Abundance of such organisms is associated with
organic pollution.
Draining of Lake Hula
and expansion of fish-breeding in the catchment would
increase influx of organic matter into L. Kinneret,
and account for the rise in lavnun biomass to a peak
in the mid-1980's. Settling ponds and reduction
of fish-breeding reversed this trend.
Organic matter influxed
through rainfall probably accounts for part of lavnun
biomass fluctuation, including the abundance of the 1991/92 brood. However, both the1990/91and the1991/92 broods were exceptionally
strong, implying an additional factor and a higher concentration of organic
matter in the water column than in previous years.
In 1990 and in 1991 there were
increases in suspended matter, total phosphorus and particulate organic
nitrogen in the water column, and a sharp increase in respiration rate on the
lake bottom (Kinneret Limnological
Laboratory reports T8/91 and T24/94). This is the eutrophic
state that Serruya & Pollingher
(1977) predicted would result from lowering water level below -212 m. Thus, the
superabundance of the 1990/91 and 1991/92 lavnun
broods was related to an increase in authochthonous
carbon resulting from the lowering of lake level to nearly -213 m (Landau,
1996; 1997).
Excepting my own articles, no
publication as far as I know mentions either organic matter or the Kinneret water level in the context of lavnun
biomass fluctuations. With the statement "An unusually successful
recruitment of lavnun during the flood winter
1991-1992 led to a sharp two- to three-fold rise in the abundance of
sub-commercial-sized fish", Ostrovsky & Walline (1999) acknowledge the effect of rainfall on the lavnun stock, but not the organic matter washed down in the
rain. Their 'sharp' abundance rise is grossly under-estimated, as can be seen
from data in this section as well as and the 'dilul' section and Landau (1996;
1997; 1998).
Collapse of the commercial lavnun stock in the 1990's was foreseen in catch-and-effort
and size-distribution data of the 1980's (Landau, 1991; 1996). Ignoring this
data, Hambright & Shapiro (1997) claim over-fishing
of large specimens in 1990 - 1993 as the cause of stock collapse. Yet they
present size distribution data that indicate recruitment failure (large
specimens only) rather than over-fishing (dominance of smaller specimens). Ostrovsky & Walline (1999)
claim starvation as the cause of stock collapse in 1993.
Because
crustacean zooplankton comprised most of the gut content of adult lavnun in the 1950's and 1970's (Komarovsky, 1952; Gophen & Landau, 1977),
it has been assumed that availability of these organisms limits the lavnun population. This assumption is challenged by the
expansion of the lavnun stock beyond the level at
which it could be sustained by zooplankton production (data in following
sub-section and in Landau, 1998)
It can be
surmised that food sources other than Cladocera and Copepoda became important in the diet of juvenile and adult
lavnun in the early 1990's. Some cyprinids are known
to be facultative feeders: they switch preference according to the relative
abundance of the food source, and thrive in eutrophic
conditions (Giussani et al, 1985, and others).
The 1995
feeding experiments of Grossart et al (1998) show that juveniles of Mirogrex and a closely related species, Acanthobrama lissneri,
were able to feed on macroscopic organic aggregates called 'lake snow' and
reject aggregates composed of cyanobacteria. From the
reduced percentage of organic carbon in fecal pellets it was estimated that the
fish digested about one third of the ingested organic carbon.
According to Grossart et al (1998), L. Kinneret
has enhanced potential of aggregate formation due to high concentrations of POC
(particulate organic matter), TEP (transparent exopolymer
particles) and cations. Experimental work has shown
that nutrients increase both the number and the size of aggregates in lake
water (Ehmann & Walters, 1998: http://www.mtsu.edu/~scientia/mainmenu/
Vol.1, issue 2).
Studying the Kinneret
food web by means of carbon isotopes, 1989 – 1992, Zohary et al (1994) claim to have confirmed the
zooplankton > Mirogrex pathway. However, their
results do not rule out organic matter as a food source, since values for POC
(particulate organic carbon) and seston, averaged
together, give the same carbon isotope value as Mirogrex.
Ecological
impact of the sardine stock
The
carbon-flow studies of Serruya et al, 1980 for Lake Kinneret
in the 1970’s describe two major pathways: Peridinium
> detritus, and 'small' phytoplankton > zooplankton > Mirogrex. Sardine biomass was relatively low then, ~5000 T,
and not considered to be of ecological importance.
The 1993
study of Walline et al purports to be an update of the
1980 carbon-flow paper, using the ECOPATH II model. This model presumes a
system in steady state, which was certainly not the case for L. Kinneret in the 1990's. No mention is made of the lowering
of the lake level and accompanying changes in the fish population. However, the
authors do present data indicating a great impact on the ecosystem of detritus,
bacteria and protozoa.
Lavnun and tilapia
population parameters given in the ECOPATH paper differ from published data (Landau, 1979, 199l; Landau et al, 1988),
which are not mentioned. Instead, the authors misquote my unpublished 1977
project report by equating the production/biomass ratio (P/B) with the
co-efficient of total mortality, Z. This is not true, and does not appear in
any of my reports or publications.
Lavnun population parameters of the ECOPATH
paper compared to parameters based on Landau, 1991. Q/B is the
consumption/biomass ratio.
__________________________________________________________________________
*Calculated
from growth rates only, spawn not included.
** According
to Walline et al, 1993 fish eat ~12% of herbivorous
zooplankton production, predatory copepods, at least 4 times as much; 90% of
zooplankton is consumed.
***Calculated
from published data for 1983-1985.
The wide
differences in biomass, P/B and Q/B are due to the suppression of data on
biomass fluctuation and growth by Walline et al.
Nevertheless, their net result in regard to food consumption is not far from my
estimate, 65% of zooplankton production, for the 15,000 - 20,000 T Mirogrex stock of the mid-1980’s. The underlying assumption
of both studies is an almost total dependence of the lavnun
on crustacean zooplankton.
In the
mid-1990's zooplankton production was lower, probably <100,000 T/a (based on
reports of the Kinneret Limnological
Laboratory), while the lavnun stock rose to 30,000 T
and required some 180,000 T/a for sustainance.
Therefore, adult lavnun must have supplemented their
diet with ‘lake snow’ and small organisms such as protozoa and rotifers. This
suggests that the cyprinid stock becomes a major cleaner of the ecosystem in eutrophic conditions (Landau, 1997; 1998).
In contrast, Gophen, (1995), Pisanti & Shapiro
(1997) and others assume a negative impact of the lavnun on water quality through its predation on
zooplankton. Undoubtedly, the lavnun has an impact on
zooplankton biomass, but other factors may be more important. Zooplankton
biomass continued to decline between 1987 and 1992, a period of decreasing lavnun
stock (Figure entitled 'Biomass trends' above). In any case, predation benefits
Kinneret water quality by cleaning and stabilizing
the ecosystem.
Possibly,
mass mortality of the sardine stock due to parasite infestation did some damage
to water quality. The 'top' predators of the Kinneret
ecosystem, the catfish Clarias lazera (safamnun) and barbels (binnit) are known to eat lavnun,
but their combined populations were insignificant in relation to the lavnun biomass in 1997. Therefore, it is likely that most
of the affected specimens died and deteriorated in the water column.
Berman et al (1998) acknowledge a
'dramatic increase in the total number of fish' without mention of water
level. Citing Hambright & Shapiro(1997) they blame the collapse of the commercial lavnun fishery in the early 1990's on the small size of
these fish. In fact the fishery had declined before the
population explosion. (further discussion in webpage
on Mirogrex biomass changes)
Home