O tecido pode ser MERISTEMÁTICO , aqueles envolvidos na formação de novas células, e PERMANENTES , aqueles nos quais formam regiões dentro da planta onde o crescimento é pelo menos temporariamente parado e nos quais células e tecidos já estão diferenciados. O crescimento em plantas não é uniformemente distribuído, mas é restrito a certas zonas contendo células recentemente produzidas pela divisão celular em um meristema.
É importante não confundir crescimento ( definido como um aumento no tamanho) com divisão celular nos meristemas. A divisão celular sozinha não causa aumento em tamanho, mas o produto da divisão celular faz crescer e causa crescimento.
Duas zonas meristemáticas principais são encontradas perto da raiz e ápices, outra no câmbio vascular, e outra justamente acima do nó das monocotiledôneas.
Os meristemas podem ser PRIMÁRIOS e SECUNDÁRIOS. Os meristemas apicais da raiz e broto são formados durante o desenvolvimento embrionário e são denominados de MERISTEMAS PRIMÁRIOS . O câmbio vascular e as zonas meristemáticas das folhas das gramineas não são distintos até depois da germinação : eles constituem os MERISTEMAS SECUNDÁRIOS.
Para elaborar seu próprio alimento através do processo da fotossintese, a planta necessita somente de água (provenienete do solo), dioxido de carbono (da atmosfera), e luz (do sol). A água juntamente com os nutrientes é absorvida nas raízes, através da epiderme, cortex, e endoderme até o tecido vascular da raiz, e move-se pela parte exterior do xilema (alburno) através do elementos xilemáticos condutores, até às folhas, onde finalmente é transpirada através do estomatos para a atmosfera. O dioxido de carbono é absorvido através de minúscula abertura na superfície foliar. Com a ajuda dos raios solares, a água é combinada na presença da clorofila para formar os açucares que fornecem energia para a árvore em crescimento. Parte dos açucares produzidos vão para locais especiais na madeira onde são armazenados para uso posterior, e parte é consumido pela respiração. Os açucares aproveitados para a produção da madeira desce através do floema.
O CÂMBIO, uma fina camada entre o xilema e floema é responsável pela produção de novo tecido de xilema e floema. desde que a seiva (solução contendo açucares, água , reguladores de crescimento e outras substâncias) desce através do floema, e é necessário no câmbio, existe a necessidade de um caminho para que possa mover-se horizontalmente em direção ao centro da árvore. Este movimento horizontal é realizado pelos RAIOS. Estes funcionam também como armazenadores de carbohidratos.
FORMAÇÃO DO TRONCO
A zona meristemática no ápice do caule principal, denominada de MERISTEMA APICAL, controla até um certo ponto o desenvolvimento dos galhos. A divisão celular neste meristema apical ocorre para alongar o tronco. A produção de novas células nesta zona meristemática é seguida de alongamento dessas células resultando desta maneira no crescimento em altura. A medida que o tronco é desenvolvido através da produção de novas células durante o período de crescimento, a gema terminal move-se para cima deixando para trás as células em expansão. Assim , se enfiarmos um prego em um tronco a 2m acima do nível do solo, este prego sempre estará a 2m do solo, independente da altura que a árvore atingir.
A produção de células no ápice do caule e o subsequente alongamento dessas células são seguidas por várias mudanças nas células maduras formadas.
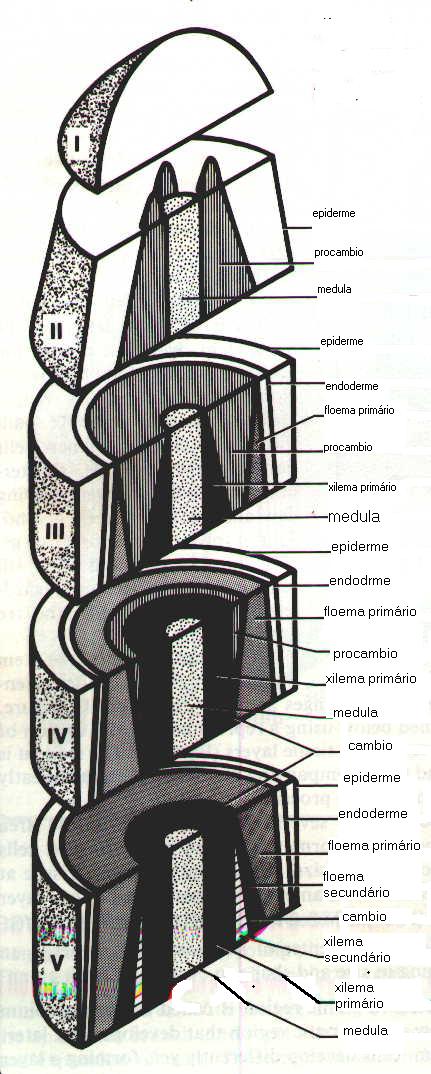
A Figura 01 mostra esquematicamente, e de uma forma simplificada a seqüência do desenvolvimento do tecido primário e secundário.
Na área I é onde novas células são produzidas, e onde está localizado o PROMERISTEMA. As células formadas sofrem mudanças em tamanho, forma e função. Na área II estão distintas três camadas diferentes: epiderme, procâmbio e medula. A epiderme é composta de uma camada de células que possuem uma parede exterior espessa envolvida pela cutina, a qual serve com proteção contra a perda de umidade. O procâmbio é um precursor de novas regiões meristemáticas que se desenvolvem posteriormente. Uma segunda camada de proteção é formada abaixo da epiderme e é denominada de endoderme ( área III). O procâmbio alcança um tamanho máximo, formando um cilindro, e as suas células mais externas e internas começam a se diferenciar em floema e xilema primário. este processo de transformação em floema e xilema primário continua até que somente uma a várias células procambiais constituem a largura total do procâmbio.
Como estágio final de um processo evolucionário que começou no ápice do caule, o tecido procambial restante torna-se ativo trasnformando-se numa camada meristemática de nominada de CÂMBIO VASCULAR , que começará a formar floema secundário ( casca mais externa) para o exterior e xilema secundário ( madeira) para o interior. Assim, todos os tecidos formados antes da formação do câmbio vascular no caule jovem são denominados de tecidos primários. O novo meristema lateral é considerado de origem secundária .
É interessante notar na maioria das mocotiledôneas todas as células do procâmbio diferenciam-se em xilema ou floema primários, e não existe a formação do câmbio vascular. Essas plantas, portanto, não produzem xilema e floema secundário. Entretanto, existem algumas monocotiledôneas que apresentam crescimento secundário derivado de atividade cambial. Quando o câmbio está presente, sua origem e ação são distintas daquelas encontradas entre as dicotiledôneas ( Philipson & Ward, 1965).Com a atividade cambial, a quantidade de floema e xilema secundários aumentam, enquanto que o floema primário é reduzido por esmagamento, e o xilema primário é depositado entre o xilema secundário e a medula.
FORMAÇÃO DA PAREDE CELULAR PRIMÁRIA
A divisão de uma célula (citocinese) inicia-se pela produção de uma placa celular, a qual origina-se pela fusão de centenas de finas vesículas. A medida que as vesículas se juntam é formada a LAMELA MÉDIA . Seguindo-se a formação desta placa celular, a formação da PAREDE PRIMÁRIA é iniciada pela deposição de hemiceluloses que polimerizam-se na parede e perdem sua solubilidade na água (Wangaard, 1970). As vesículas de Golgi no protoplasma são possivelmente um importante fator nesta seqüência da formação a parede primária, podendo atuarem como sintetizadores dos polissacarídeos não celulósicos ( Salisbury & Ross, 1985).
Assim, a parede primária das células em crescimento consiste largamente de uma matriz de polissacarídeos não celulósicos e algumas proteínas, onde existem microfibrilas celulósicas. Essas microfibrilas podem ser sintetizadas através da agregação de cadeias moleculares de celulose, ou desenvolver-se no fim da síntese envolvendo a transferência de glucose ou pequenos polímeros solúveis na água sucessivamente para o fim da cadeia já incorporada nas micrifibrilas (Preston, 1962). No último processo poderia haver um aumento no comprimento das microfibrilas. Para que isso ocorra, polimerização e cristalização devem ser eventos essencialmente simultâneos.
É importante sabermos o que causa o alongamento das células na maior parte das vezes em uma direção ao invés de se expandirem igulamente em todas as direções. Conforme comentado anteriormente a parede primária das células em crescimento consistem largamente de uma matrix amorfa de polissacarídeos não celulósicos e algumas proteínas, através das quais existem microfibrilas de celulose que minimizam o alongamento na direção axial longitudinal. As microfibrilas não têm a propriedade de se alongarem facilmente.
Assim , a formação da parede celular pode ocorrer efetivamente na direção que permite muitas microfibrilas "deslizarem" entre si. O crescimento é favorecido na direção que forma um ângulo reto com o eixo longitudinal das microfibrilas. Além disso , quando novas moléculas de celulose são formadas durante o crescimento, as microfibrilas celulósicas existentes são aparentemente alongadas permitindo uma extensão paralela ao seu eixo ( Salisbury & Ross, 1985).
Em algumas células jovens, a orientação das microfibrilas não é ao acaso, de maneira que o crescimento começa acelerado ao longo de um eixo (Jansen & Salisbury, 1984). A medida que o crescimento contínua, novas microfibrilas são depositadas na parede adjacente à membrana plasmática de maneira que a parede retem uma espessura uniforme durante o crescimento. Se a orientação dessas microfibrilas é ao acaso, o crescimento tende a ser igual em todas as direções; mas se são depositadas mais perpendicular a um eixo, o crescimento é favorecido na direção daquela orientação (Figura 2) (Salisbury & Ross, 1985).
FORMAÇÃO DA PAREDE SECUNDÁRIA
pós o desenvolvimento da parede primária é a fase do espessamento da parede ou formação da parede secundária. A formação dessa parede secundária começa próximo ao meio de uma fibra e prossegue em direção às extremidades onde o alongamento da fibra ( e portanto expansão da parede primária) pode estar ainda estar ocorrendo (Wardrop, 1964). O desenvolvimento de sucessivas lamelas da camada S1 da parede secundária é representada diagramaticamanete na Figura 03. Devido as direções opostas da orientação hel;ical nas sucessivas lamelas de S1,segue-se que asa hélices microfibrilares do tipo Z e S são colocadas simultaneamente em diferentes partes da mesma célula.
A orientação nas numerosas lamelas da camada S2 da parede secundária tem sido mátéria de estudo de muitos investigadores. A orientação microfibrilar da primeira lamela na parede secundária é pouco diferente daquela na parede primária.
[ Identificando ] [ Macroscopia ] [ Cerne ] [ Folhosas X Coniferas ] [ Variabilidade dos .........]