�H������www.oocities.org/es����������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������/tonicasany/apuntscell_3.html���������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������www.oocities.org/es/tonicasany/apuntscell_3.html�elayed������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������x������J����������������������������������������������������������������������q�����������������������������OK������������������������������������������������������������������������������text/html����������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������O0`�����������������������b�.H��������������������������������������������������������������������������������������������������������������������������������������������������������Wed, 14 Feb 2007 17:54:54 GMT�������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������Mozilla/4.5 (compatible; HTTrack 3.0x; Windows 98)��������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������en, *����������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������J����������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
membrana i paret cel·lular
2. Estructura de la cèl·lula eucariota
2.1. La membrana plasmàtica
2.1.1.
Composició i estructura de la membrana plasmàtica
2.1.2.
Funcions de la membrana plasmàtica
2.1.2.1.
Permeabilitat cel·lular
2.1.2.1.1. Transport
de molècules petites
2.1.2.1.1.1
Transport passiu
2.1.2.1.1.2
Transport actiu
2.1.2.1.2.
Transport de macromolècules
2.1.3.
Diferenciacions cel·lulars
2.2.
Cobertes i parets cel·lulars
2.2.1.
Glucocalix
2.2.2.
Paret cel·lular de les cèl·lules vegetals
COMPONENTS DE
LA CÈL·LULA EUCARIOTICA

Rough ER= reticle endoplàsmic rugós Smooth ER= reticle endopàsmic llis
Animació de John Kyrk sobre la cèl·lula (en anglés)
|
COMPONENT |
ESTRUCTURA
|
FUNCIÓ
|
|
Membrana
cel·lular
|
Estructura
en mosaic fluíd: bicapa lipídica amb proteïnes
( +glucocàlix extern en cèl·lulas animals)
|
Límit de la cél·lula
i permeabilitat selectiva
|
|
Paret
cel·lular
|
Paret primària i paret
secundària de fibres de cel·lulosa
|
Responsable de la forma de las
cèl·lules; les atorga suport mecànic i manté
l'equilibri osmòtic
|
|
Hialoplasma
|
Solució
aquosa amb alta concentració de proteïnes, principalment
enzimàtiques
|
Participació
en processos metabòlics
|
|
Citoesquelet
|
Xarxa tridimensional formada per filaments proteics.
|
Organització i control
de l'espai interior. Involucrat en la forma, moviment i divisió
cel·lular.
|
|
Centríols
|
Microtúbuls
i microfilaments proteics
|
Formació del fus (mitòtic) acromàtic.
Formació de cil·lis i flagells.
|
|
Ribosomas
|
Dues subunitats formades per ARN i proteïnes
|
Síntesi
de proteïnes
|
|
Reticle
Endoplàsmic Rugós
|
Cisternes membranals intercomunicades
i amb ribosomes adherits
|
Síntesi,
processament i emmagatzegament de proteïnes
|
|
Reticle
Endoplàsmic Llis
|
Cisternes de membrana intercomunicades
|
Síntesi,
emmagatzegament i transport de lípids. eliminació
de substàncies tòxiques.
|
|
Aparell
de Golgi
|
Sistema de cisternes de membrana aplanades
|
Maduració,
emmagatzegament i transferència de glucoproteïnes.
Formació de membranes, i paret cel·lular.
|
|
Lisosomes
|
Vesícules
esfériques de membrana que contenen enzims digestius.
|
Digestió
cel·lular
|
|
Peroxisomes
|
Vesícules esfériques
de membrana que contenen enzims oxidatius
|
Protecció contra productes
tòxics del metabolisme
|
|
Vàcuols
|
Vesícules arrodonides
|
Emmagatzemen
substàncies: aigua, nutrients, substàncies de rebuig.
|
|
Mitocondris
|
Orgànuls amb doble membrana.
Presenten gran quantitat d'enzims, ADN i ribosomes
|
Centrals energètiques
de la cèl·lula: duen a terme la respiració
cel·lular, consistent en l'oxidació de nutrients
per a obtenir ATP.
|
|
Cloroplasts
|
Orgànuls amb doble membrana
més una tercera en el seu interior (tilacoidal). Conté
enzims, pigments fotosintètics, ADN i ribosomas.
|
Responsable de la fotosíntesi.
|
|
Membrana
nuclear
|
Doble
membrana amb porus
|
Separar i protegir l'ADN de
la resta de la cèl·lula
|
|
Nucleoplasma
|
Composició
semblant a l'hialoplasma
|
.Conté
enzims involucrats en la replicació de l'ADN i en la transcripció
de l'ARN
|
|
Cromatina
|
ADN més proteïnes densament empaquetat
|
Portador
de la informació genètica
|
|
Nucléol
|
Regió
esferoidal amb alta concentració d'ARN i proteïnes
|
Lloc
de síntesi de les subunidats ribosòmiques.
|
Tornar
a l'inici de la pàgina
2.1.
Membrana plasmàtica
L'aparició de les
membranes plasmàtiques va constituir un fet fonamental
en l'origen dels primeres formes de vida. La membrana plasmàtica,
que envolta totes les cèl·lules, defineix l'extensió
d'aquesta i manté les diferències essencials entre
el seu contingut i el del medi extern. La membrana és, però,
qualque cosa més que una simple barrera passiva: és
un filtre altament selectiu que manté la desigual concentració
d'ions a l'exterior i l'interior de la cèl·lula
i permet l'entrada de nutrients a la cèl·lula, així
com l'eixida de productes residuals. Totes les membranes biològiques,
incloses la membrana plasmàtica i les membranes
internes de les cèl·lules eucariotes presenten
una estructura comuna: una agrupació de molècules
lipídiques i proteïnes unides per interacions no covalents.

Relació superficial entre la membrana plasmàtica, el conjunt de membranes internes i el citosquelet. La cèl·lula no és eixe globus ple d'aigua que us havien dit fins ara

Segons el model de mosaic fluïd desenvolupar per Singer
i Nicolson (1972) les molècules lipídiques
es presenten en forma de una doble capa contínua (bicapa
lipídica) que constitueix l'estructura bàsica
de la membrana i actua de barrera relativament impermeable al pas de la majoria de molècules hidrosolubles (aquesta
impermeabilitat és relativa ja que s'ha desenvolupat tota
una sèrie de sistemes de transport de substàncies
a través de membrana). Les molècules de proteïnes
es trobem inmerses en la bicapa lipídica. Totes les membranes
cel·lulars són estructures fluïdes (Veure animació),
flexibles, dinàmiques: la majoria de les seues molècules
lipídiques i proteiques poden desplaçar-se pel pla
de la membrana (no hi ha enllaços covalents entre els fosfolípids
ni entre fosfolípids i proteïnes sinó que tota
l'estructura de la bicapa es manté per l'acció d'enllaços
dèbils). A més a més, són estructures asimètriques ja que la composició de les
dues cares és diferent com a resposta de les diferents
funcions realitzades per les dues superfícies.

Fig.15: Components i estructura
de mosaic fluid de la membrana plasmàtica

Fig.16: Microfotografia al MET
la membrana plasmàtic de dues cèl·lules adjacents
Components
de la membrana

Lípids de membrana
* La bicapa lipídica és fluïda i les diferents molècules lipídiques
poden difondre ràpidament dintre de la pròpia monocapa,
mentre que rarament passen d'una monocapa a una altra. Les molècules
lipídiques de la membrana són amfipàtiques,
i la major part d'elles formen espontàniament bicapes quan
estan en contacte amb l'aigua. Per aquesta raó les bicapes
lipídiques cel·lulars es formen per auto-acoblament
i quan es trenquen, s'uneixen novament (autosegellament).
A la bicapa de la membrana plasmàtica hi ha tres tipus
principals de molècules lipídiques - fosfolípids,
colesterol i glicolípids -Les diferents membranes d'una
cèl·lula eucariota presenten composicions lipídiques
diferents; les següents dades pertanyen a la membrana plasmàtica
Els fosfolípids són les molècules
lipídiques més abundants i constitueixen la base
de la bicapa. Presenten la seua part polar en contacte amb el
medi aquós mentre que les cues apolars corresponents a
les cadenes hidrocarbonades dels àcids grassos romanen
a l'interior de la bicapa. El colesterol augmenta la resistència
de la membrana ja que s'intercala entre els fosfolípids
i tendeix a estalilitzar les seues cues. Els glucolípids (fosfoesfingolípids, gangliòsids, etc) es troben
a la superfície externa de la membrana plasmàtica
on constitueixen un 5% de la monocapa externa. Encara no
se sap massa sobre la seua funció però se suposa que tenen un important paper en funcions de reconeixemen cel·lular.

Fig 17. Disposició
dels fosfolípids dins de l'estructura de la bicapa lipídica.
Les zones polars es disposen a l'exterior en contacte amb les
molècules d'aigua, mentre que les cadenes hidrocarbonatades (apolars)
dels àcids grassos se situen a l'interior fora de contacte
amb l'ambient aquós.
 Fig.18. El colesterol estabilitza
la bicapa lipídica
Fig.18. El colesterol estabilitza
la bicapa lipídica
Proteïnes de membrana
* Si la bicapa lipídica determina l'estructura bàsica
de les membranes biològiques, les proteïnes són les responsables de la majoria de les seues funcions,
ja que actuen com a receptors, enzims o transportadors específics.
Moltes proteïnes posseeixen superfícies hidròfobes
que les fan insolubles en una solució aquosa i s'associen
a la bicapa lipídica de diferents formes: moltes s'estenen
a través de la bicapa (proteïnes transmembranals)
amb els radicals hidròfobs en contacte amb les cues apolars
dels fosfolípids i els radicals polars situats a ambdós
costats de la bicapa lípidica; d'altres proteïnes
s'indrodueixen parcialment en la bicapa lipídica mentre
que tenen una altra part exposada al medi aquós; finalment
hi trobem proteïnes situades fora de la bicapa, unides a
proteïnes transmembranals o lípids. A l'igual que
les molècules lipídiques moltes de les proteïnes
membranals poden difondre en el pla de la membrana.
* A la membrana plasmàtica de totes les cèl·lules
eucariotes, la majoria de les proteïnes que resten exposades
sobre la superfície cel·lular i algunes de les molècules
lipídiques de la monocapa lipídica externa es troben
unides covalentment a cadenes d'oligosacàrids. Encara
que no es coneix la funció d'aquests hidrats de carboni,
sembla probable que participen en fenòmens de reconeixement
cèl·lula-cèl·lula. (veure imatge)

 Fig. 20. Tipus
de proteïnes de membrana
Fig. 20. Tipus
de proteïnes de membrana
Animació de John Kyrk sobre la membrana plasmàtica. Els diferents tipus de lípids que la formen, proteïnes de membrana, etc. Animació molt bona, tot i que costar captar la totalitat de la informaó que proporciona
Tornar
a l'inici de la pàgina
2.1.2.Funcions de la membrana
-
Mantindre les
diferències de composició existents entre el medi cel·lular i
el medi extern. També actuen com a barrera que permet
la cèl·lula mantenir unes condicions físico-químiques
particulars a l'interior de l'orgànul que delimiten (nucli,
mitocondri, etc): delimitació de compartiments cel·lulars
-
Mantenir
una permeabilitat selectiva per mitjà del control
del pas de substàncies entre l'exterior i l'interior.(veure
la següent pregunta)
-
Produir, modular i conservar gradients electroquímics
entre un costat i un altre de la membrana. El manteniment
d'aquest gradient electroquímic a banda i banda de la
membrana plasmàtica constitueix el fonament físico-químic
de l'excitabilitat, la propietat que tenen les cèl·lules
de reaccionar front els estímuls de l'ambient i elaborar
respostes adequades. Veure l'animació
-
Rebre i transmetre senyals. A la membrana plasmàtica
s'hi troben sistemes encarregats de rebre informació de
l'ambient. Aquests sistemes consten fonamentalment de receptors
de senyals localitzats a la superfície externa de la membrana
i mecanismes de transducció de senyal localitzats a la
superfície interna i que transformen un esdeveniment extracel·lular
en un senyal intracel·lular. En els organismes pluricel·lulars
les cèl·lules es comuniquen entre elles a través
de molècules que actuen com a missatgers químics
de la informació (hormones, neurotransmissors, etc). Es
tracta de substàncies alliberades per grups de cèl·lules
especialitzades que travessen l'organisme per actuar sobre determinades
cèl·lules diana; és a dir, una molècula
missatge determinada únicament obté resposta d'aquelles
cèl·lules que tenen a les seues membranes receptors
adequats. Posteriorment els mecanismes de transducció
provocaran l' aparició dins la cèl·lula
d'una altra molècula - segon missatger- que hi
exercirà l'acció : AMPc
 Un mecanisme típic de senyalització cel·lular. L'arribada d'una molècula senyalitzadora (com per exemple una hormona) i el seu encaix amb un receptor de membrana provoca tota una sèrie de canvis a nivell de la membrana que té com a resultat la produccó i alliberament de determinades subsàncies que viatjaran fins al nucli on actuaran en la regulacó de la transcripció. Si vols veure una animacó d'aquesta imatge, punxa ací
Un mecanisme típic de senyalització cel·lular. L'arribada d'una molècula senyalitzadora (com per exemple una hormona) i el seu encaix amb un receptor de membrana provoca tota una sèrie de canvis a nivell de la membrana que té com a resultat la produccó i alliberament de determinades subsàncies que viatjaran fins al nucli on actuaran en la regulacó de la transcripció. Si vols veure una animacó d'aquesta imatge, punxa ací
Tornar
a l'inici de la pàgina
2.1.2.1. PERMEABILITAT CEL·LULAR
2.1.2.1.1 Transport
de molècules petites a través de la membrana
La bicapa lipídica,
com a conseqüència de la seua estructura, actua com
a una barrera altament impermeable a la majoria de les molècules
polars, tot impedint que la major par del contingut cel·lular
-hidrosoluble- es difonga cap a l'exterior. Tanmateix, les cèl·lules
han d'obtenir del medi exterior tota una sèrie de nutrients
polars així com desfer-se de'ions i molècules orgàniques.
Aquesta és la causa de l'aparició de perfeccionats
sistemes de transport a través de la membrana.
La permeabilitat de substàncies a través de la bicapa
lipídica depén:
* Grandària (quan més xicotetes millor)
* Caracter polar (passen millor les molècules apolars,
sense càrregues)
* Gradient de concentració
Si estudiem el comportament de diferents molècules a través
de bicapes lipídiques artificials obtenim els següents
resultats:
|
Molècules
hidrofòbiques (no
polars, sense càrregues elèctriques)
|
O2, CO2, bencè
|
Es
dissolen ràpidament en la bicapa i hi difonen
|
|
Petites
molècules polars sense càrrega
|
aigua, urea, glicerina
|
Es
dissolen en la bicapa i hi difonen
|
|
Molècules grans sense
càrrega
|
glucosa, sacarosa
|
No poden travessar la bicapa
|
|
Ions
|
H+, Na+, Cl-
|
Les bicapes són altament
impermeables a totes les molècules carregades
|
A l'igual que les bicapes lipídiques artificials, les membranes
biològiques permeten el pas de l'aigua i de molècules
no polars per simple difusió, tot i que també són
permeables a diverses molècules polars com ions , monosacàrids
, aminoàcids , etc que travessen amb una gran lentitud
les bicapes lipídiques artificials. Això és
possible gràcies a l'existència de diferents tipus
de proteïnes de membrana que són les responsables
del transport de substàncies específiques. Algunes
proteïnes de transport simplement transporten un solut d'una
part a l'altra de les membranes (transport senzill o uniport).
D'altres funcionen com a sistemes de cotransport, on la transferència
d'un solut depén de la transferència d'un altre
compost, ja siga en el mateix sentit (simport) o en el sentit
oposat (antiport)

Fig.21: Tipus de transport en
funció de les substàncies que hi participen
Si atenem a raons energètiques
podem distingir altres dos tipus de transport a través
de membrana: Transport passiu i transport actiu
Tornar
a l'inici de la pàgina
2.1.2.1.1.1.Transport passiu.
Es tracta d'un procés espontani que té lloc sense
despesa d'energia metabòlica (ATP) per part de la cèl·lula.
La direcció del transport ve governada pel:
- Gradient de concentració (molècules sense càrrega)
- Gradient electroquímic (concentració i gradient
elèctric)
Normalment les cèl·lules tenen un potencial de membrana
negatiu - són més negatives a l'interior- fet que
facilita l'entrada d'ions positius i s'oposa a l'entrada d'anions.
Pot tenir lloc mitjançant:
- Osmosi: la cèl·lula intercamvia aigua amb
el medi extracel·lular. (Veure l'animació)
- Difusió a través de bicapa. La cèl·lula
intercamvia molècules dissoltes per massa reduïda
i de caracter apolar a través de la bicapa lipídica.
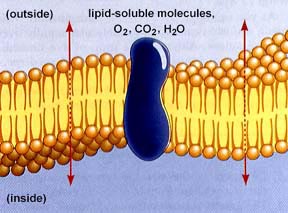 Fig.22: Difusió a través
de bicapa
Fig.22: Difusió a través
de bicapa


Fig.23: Transport
a través d'una proteïna de canal regulat per lligand
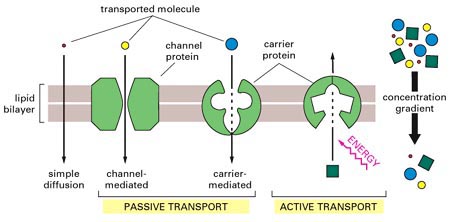
Fig.24: Diferents
tipus de transport passiu i actiu
Tornar
a l'inici de la pàgina
2.1.2.1.1.2 Transport actiu
Aquest tipus de transport requereix una despesa d'energia metabòlica
en forma d'ATP. En aquest cas, les proteìnes transportadores
poden transportar molècules contra un gradient de concentració
o electroquímic. Aixó implica la hidròlisi
d'ATP o un cotransport a favor dels seus gradients electroquímics
Les proteines transportadores es comporten ací com a enzims
(unió específica amb la molècula per transportar).
Sovint sofreixen canvis de conformació reversibles quan
passen molècules a través de la membrana. En ocasions solament hi ha un únic tipus de molècula (o ió) transportada (uniporte, veure l'animació) però també és possible aprofitar la despesa energètica per fer passar una altra molècula a través de membrana: quan els dos tipus de substàncies viatgen en un mateix sentit parlem de simporte (veure animació) i quan ho fan en sentit oposat, antiporte (veure l'animació)

Fig.25: Tipus de transport
actiu
Un exemple que il·lustra el mecanisme de transport actiu
és l'anomenada bomba Na+ / K+ (veure
l'animació fent clic) present a totes les cèl·lules
animals. Aquest mecanisme de transport actiu funciona com un transport
d'intercanvi (antiport) ja que bombardeja cap a l'exterior ions
sodi i cap a l'interior ions potassi en contra dels seus gradients
de concentració (en el cas del Na+ també en contra
del seu gradient elèctric)
Aquesta bomba sodi-potassi manté el potencial de membrana
(normalment les cèl·lules són més
negatives a l'interior que a l'exterior), així com participa
en el transport actiu a través de membrana de molècules
com aminoàcids i glucosa. La cèl·lula gasta
1/3 del total de la seua energia en mantindre aquest mecanisme
de transport (un 70% a les neurones).
L'energia necessària per a què tinga lloc aquest
tipus de transport prové de la hidròlisi de l'ATP
catalitzada per una proteïna l'ATPasa, enzim al·lostèric
que presenta un canvi de conformació que afavorirà
el pas de les molècules a través de la membrana

Fig.27. Un exemple de
transport actiu: la bomba sodi-potassi. Es tracta d'un exemple
d'antiport on la cèl·lula fa entrar 2 ions de potassi
al mateix temps que fa eixir 3 de sodi. Aquest transport és
dur a terme gràcies a l'alliberament d'energia per part
de l'ATP
Animació de la bomba sodi-potassi. Transport actiu antiporte
Altra animació en anglés (text i audio) sobre la bomba sodi-potassi de l'editorial McGraw Hill
Aquesta bomba sodi-potassi crea un potencial de membrana tant elèctric com de concentració: la part interna de la membrana queda negativa respecte a l'externa, ja que el nombre de sodis i potassis que surten i entren són diferents. però també crea un important gradient de concentració del sodi, que va acumulant-se a l'exterior i té una tendència natural a tornar a entrar /a favor de gradient elèctric i de concentració). Això ho aprofita la cèl·lula per fer-lo passar per una proteïna transportadora especial i per fer que cada vegada que el sodi torne a entrar a la èl·lula ho faça acompanyat d'una molècula de glucosa (en contra de gradient de concentració) que aprofitara l'entrada del sodi, per entrar en la cèl·lula sense necessitat de despesa energètica. Tot aiò queda més clar amb l'animació següent.
Continua......
Index
general de la cèl·lula
{kind=link}
{kind=link}
{kind=link}