�H������www.oocities.org/es����������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������/tonicasany/apuntscell_5.html���������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������www.oocities.org/es/tonicasany/apuntscell_5.html�elayed������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������x������J����������������������������������������������������������������������q����������������������e������OK������������������������������������������������������������������������������text/html���������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������`*1`����e������������������b�.H��������������������������������������������������������������������������������������������������������������������������������������������������������Wed, 14 Feb 2007 18:48:23 GMT�������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������Mozilla/4.5 (compatible; HTTrack 3.0x; Windows 98)��������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������en, *����������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������J���������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������e������
Índex cèl·lula
Citologia i fisiologia cel·lular. El citoplasma(II): orgànuls membranosos
1. Reticle endoplasmàtic
(RE)
El RE és un orgànul membranós de totes les cèl·lules eucariotes format per un
conjunt de membranes que constitueix més de la meitat de
la membrana total de la cèl·lula; aquest sistema
membranós delimita un espai intern anomenat lumen del RE el qual apareix fragmentat en forma de cavitats
planes interconnectades o bé en forma de cisternes, túbuls
i vesícules, tot i que es creu que hi ha una única
làmina membranosa que envolta el lumen, com si fora un
sac replegat diverses vegades.
Hi ha una zona del RE que conté ribosomes associats a la
cara externa de la seua membrana i pel seu aspecte al microscopi
electrònic rep el nom de RE rugós (RER),
mentre que la zona que no té ribosomes s'anomena RE
llis (REL). Aquestes dos regions també difereixen pel
que fa a la seua forma: el RER està organitzat en piles
de sàculs aplanats mentre que el REL està constituït
per una sèrie de túbuls fins, molt sovint ramificats.
Una part del RE rugós continua amb la membrana externa
del nucli mentre que una altra part es comunica dinàmicament
amb l'aparell de Golgi, a través de vesícules transportadores
.

Fig.1: Diferències estructurals entre els dos tipus de
RE: RER a l'esquerra i REL a la dreta

Fig.2: Fotografia amb MET (microscopi electònic de transmissió) del RER (sàculs aplanats de color violeta) i els ribosomes adosats a la
seua superfície (punts de color verd)

Fig.3: Esquema que mostra les relacions estructurals del RE i l'embolcall nuclear. Les escructures del RER es troben disposades concentricament al volrant del nucli i existeir una continuïtat entre la membrana externa de l'embolcall nuclear i les membranes del RER
Funcions
Al RER, els ribosomes
adosats a la seua superfície són els encarregats
de realitzar la síntesi proteica. Part d'aquestes proteïnes
poden quedar fixades a la membrana del RE on posteriorment poden
ser exportades per formar les membranes d'altres orgànuls
cel·lulars, però la major part passarà al
lumen del RE on seran transformades per l'adició d'oligosacàrids
(glicosilació) i posteriorment seran exportades
a altres compartiments cel·lulars (Golgi) o secretades
a l'exterior.

Fig.4: Esquema que mostra les transformacions que pateixen les proteïnes sintetitzades en els ribosomes adosat a la membrana del RER en l'interior d'aquest orgànul. Aquesta "maduració " de les proteínes consisteix en l'adicó de residus glucídics
El REL -reticle endoplàsmic llis-no participa en la síntesi proteica. Malgrat
que és abundant en determinades cèl·lules (hepatòcits,
cèl·lules secretores) normalment constitueix una
petita part del RE total .
- A la membrana del REL es fa la síntesi
de diverses substàncies lipídiques com el colesterol
(i els seus derivats ) i els fosfolípids que formen part
de les bicapes de totes les membranes de la cèl·lula. També sintetitzen (en determinades cèl·lules) hormones esteroides
- També hi tenen lloc reaccions de destoxificació
que actuen sobre molècules liposolubles estranyes a l'organisme amb
la finalitat d'augmentar la seua hidrosolubilitat i accelerar-ne
l'eliminació (medicaments, drogues, etc).
- Les cèl·lules musculars tenen
un REL especialitzat anomenat reticle sarcoplasmàtic que
acumula ions Ca2+, necessaris pel procés de contracció
muscular
2. L'aparell o complex de Golgi
L'aparell de Golgi està format
per conjunts de sacs aplanats de forma discoïdal envoltats
d'un gran nombre de menudes vesícules sorgides per gemmació
dels marges dilatats de les cisternes. Està situat prop
del nucli i, a les cèl·lules animals, envolta el
centríol. Cada conjunt de sàculs rep el nom de dictiosoma.
El nombre de dictiosomes varia d'una cèl·lula a
una altra i és més gran en les cèl·lules
especialitzades en la secreció. L'aparell de Golgi presenta una polaritat, és a dir, es poden diferenciar dues cares en els dictiosomes - cis i trans- que presenten diferent composició i funció

Fig.5. Estructura polaritzada de l'aparell de Golgi
L'aparell de Golgi es troba polaritzat estructuralment i bioquímica:
presenta dos cares, la cara cis o de formació i
la cara trans o de maduració. La cara cis es localitza
prop de les membranes del RE; les seues membranes són molt
fines i de composició semblant a les del RE. Al voltant
d'ella es troben les vesícules de Golgi o de transició que deriven del RE. La cara trans es troba més sovint a
prop de la membrana plasmàtica; les seues membranes són
més gruixudes i semblants a la plasmàtica. En aquesta
cara es localitzen unes vesícules més grans, les vesícules secretores.En la cara trans també
s'han trobat enzims lisosòmics, fet que indica la seua
relació amb els lisosomes.
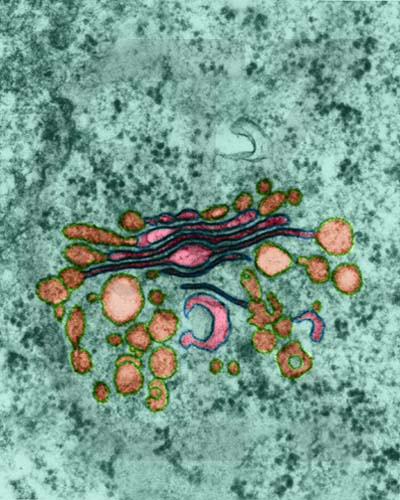
Fig.6: . Imatge
amb MET de l'estructura de l'aparell de Golgi. tall transversal de l'estructura amb color fucsia. D'aquesta estructura es desprenen/fusionen vesícules (color marró). el citosil està pintat de verd

Fig.7: Esquema de les relacions existents entre els principals orgànuls membranosos del citoplasma: 1- Embolcall nuclear; 2- Porus nuclear; 3. RER; 4-REL; 5-Ribosoma; 6-Proteïnes; 7- Vesícules de transició; 8- Aparell de Golgi; 9-Cara cis; 10-Cara trans; 11-Dictiosoma
Animació de John Kyrk sobre l'aparell de Golgi i les seues relacions dinàmiques amb altres estructures membranoses de la cèl·lula
Aquestes relacions entre els diferents orgànuls membranosos es pot demostrar mitjançant la utilització de molècules marcades radiactivament que van canviant la seua posició intracel·lular amb el pas del temps, tal com podeu observar en la figura de la pàgina 127 del vostre llibre
Funcions
- Modificació de la fracció glucídica de les glicoproteïnes i glicolípids sintetitzats
al RE. Maduració de les proteïnes
- Secreció dels productes
sintetitzats al RE. L'aparell de Golgi és el principal
director de la circulació macromolecular per l'interior
de la cèl·lula: diferents tipus de molècules
passen a través d'alguna porció del complex de Golgi
en algun moment de la seua maduració, generalment poc després
de la seua síntesi al RE (proteïnes i glucoproteïnes
que seran segregades; glucolípids i glicoproteïnes
de la membrana plasmàtica, proteïnes dels lisosomes
i la paret cel·lular). Aquestes molècules s'incorporen
a les cisternes de l'aparell de Golgi a través de les vesícules
de transició originades per gemmació de les membranes
del RE; després emigraran cap a la cara trans on passaran
a les vesicules de secreció formades per la fragmentació
de les cisternes, les quals es dirigiran a la membrana plasmàtica
on lliuren el seu contingut mitjançant un procés
d'exocitosi. En algunes ocasions (cèl·lules glandulars) aquestes vesícules es troben folrades d'unes proteïnes específiques (clatina) i reben el nom de vesícules revestides. això farà que la seua aparició estiga lligada a l'arribada d'un estímul específic
- Reciclatge de la membrana plasmàtica. La fusió
de les vesícules de secreció procedents dels dictiosomes
amb la membrana plasmàticas contribueix a la regeneració
d'aquesta
- Síntesi dels components de la paret de les cèl·lules
vegetals : cel·lulosa, hemicel·lulosa, pectina,
etc. D'ací el seu nom de membrana de secreció.
- Intervé en la formació dels lisosomes
tornar
a l'inici de la pàgina
3. Lisosomes
Són compartiments del citoplasma
que contenen enzims hidrolases adaptades a catalitzar en medi àcid
(pH 3-6) la ruptura o lisi de diferents tipus d'enllaços(peptídics, glucosídics, esters, etc). La membrana
dels lisosomes és resistent a l'acció d'aquests
enzims.
Aquests orgànuls es poden considerar en conjunt com el
estómac de la cèl·lula ja que al seu interior
es realitza la digestió de les proteïnes, hidrats
de carboni, lípids i àcids nucleics.Es poden distingir
dos classes de lisosomes: primaris i secundaris.
- Lisosomes primaris, acabats de formar i que provenen
de petites vesícules carregades d'enzims hidrolítics,
sintetitzats en primer lloc al RER i que més tard completen
la seua síntesi a l'aparell de Golgi i que s'originen per
gemmació a partir de la cara trans de les cisternesl'aparell
de Golgi.
- Lisosomes secundaris que resulten de la fusió
repetida dels lisosomes primaris amb diversos sustrats units a
membranes. Hi podem distingir els vàcuols fagocítics que provenen de la fagocitosi de grans partícules o les vesícules autofàgiques que són estructures
lisosomiques que contenen (i digereixen) membranes o orgànuls
intracel·lulars

Fig.7: Esquema que mostra la realció dels lisosomes, l'aparell de Golgi i la membrana plasmàtica, així com els processos de endocitosi, exocitosi, digestó cel·lular

Fig 9 : Tipus de lisosomes. Procés
de digestió intracel·lular: autofàgia i heterofàgia

Fig. 10. Funcions dels lisosomes
Funcions
*Els lisosomes són
els responsables de la digestió cel·lular. Els lisosomes poden digerir macromolècules d'origen extern
i intern. Aquesta funció pot realitzar-se de dos maneres
- Digestió extracel·lular: els lisosomes vessen
els seu contingut a l'exterior de la cèl·lula on
té lloc la digestió; aquest tipusd e digestió
és típica dels fongs.
- Digestió intracel·lular:
l'acció digestiva té lloc a l'interior del lisosoma.
Aquesta digestió intracel·lular es fa per heterofàgia
o per autofàgia, segons la procedència del material
que serà hidrolitzat enzimàticament
- Heterofàgia: és un procés caracteritzat
per la digestió de sustrats d'origen exogen. La seua finalitat
és doble: nutrir i defendre la cèl·lula front
a la presència de substàncies estranyes. Els substrats
són capturats per endocitosi i formen un vàcuol
endocític que es fusiona amb un lisososma primari i
originen un lisososma secundari (vàcuol digestiu).
És a l'interior d'aquesta estructura on té lloc
la digestió dels substrats. Els productes d'aquesta digestió
passen a través de membrana a l'hialoplasma on són
reutilitzats; els productes no digeribles formen un cos residual que pot fondre's amb la membranan plasmàtica i vessar eixos
productes a l'exterior per exocitosi.
- Autofàgia. En aquest cas la cèl·lula
digereix part del seu contingut citoplasmàtic, especialment
els orgànuls envellits que penetren al lisosoma par una
invaginació de la seua membrana. Aquest procés d'autodestrucció
controlada permet el recanvi continu de tots els components cel·lulars,
excepte l'ADN, gràcies a l'equilibri que mantenen les cèl·lules
entre destrucció i síntesi. L'autofàgia també
està relacionada amb la destrucció controlada dels
teixits larvaris durant el procés de metamorfosi, o amb
la reabsorció de protoplasma que fan determinades cèl·lules
com els glòbuls vermells quan es carreguen d'hemoglobina.
També hi ha autofàgia quan les cèl·lules
se sotmeten a un dejuni perllongat.
Una animació de fagocitosi
A contínuació teniu un enllaç a la pàgina de john Kyrk que ens presenta una magnífica animacó (en anglés) que resumeix les relacions funcionals entre els diferents orànuls cel·lulars (nucli, ribosomes, reticle endoplàsmic, aparell de Golgi, membrana plasmàtica) i fenòmens associats: síntesi de proteïnes , transformacó d'aquestes proteïnes en l'interir de l'aparell de Golgi ni el reticle endoplàsmic, endocitosi, exocitosi
4. Peroxisomes i gliosomes
Aquests orgànuls
reben el nom de microsomes i són un conjunt de orgànuls
esfèrics envoltats d'una membrana que allotja a l'interior
una matriu densa i compacta on apareixen sovint inclusions d'estructura
cristal·lina.
Els peroxisomes contenen al seu interior enzims oxidatius
L'enzim més abundant és la catalasa. Aquest
enzim utilitza el peròxid d'hidrogen (aigua oxigenada),
obtingut per l'oxidació de diferents sustrats per oxidar-ne
uns altres
REs reaccions oxidatices poden provocar substàncies oxidants, potencialment perilloses per a la cèl·lula com l'aigua oxigenada
R-H2 + O2 4444 R + H2O2
Aquest excés de substàncies oxidants es pot controlar per l'acció d'enzims com la catalasa que es troba a l'interior dels peroxisomes
H2O2 + X-H2 4444 2H2O + X
Aquesta darrera reacció és considerada com a un
dispositiu de seguretat que impideix una acumulació perillosa
de H2O2, que es comporta com un fort agent oxidant.
Els peroxisomes són molt abundants a les cèl·lules
hepàtiques i renals i on tenen una important funció
destoxificant ja que allí s'oxida a acetaldehid gairebé
la meitat de l'etanol que bevem. També catalitzen la degradació
d'una quarta part dels àcids grassos a acetil-CoA (la
resta al mitocondri).
Alguns autors consideren els peroxisomes com un mecanisme primitiu
(anterior als mitocondris) que va permetre les cèl·lules
eucariotes ancestrals adaptar-se a l'oxigen

Fig 11; Microfotografia d'una cèl·lula on podem diferenciar un peroxisoma, 2 mitocondris i un cloroplast
Els glioxisomes són una classe de peroxisomes anomenats
així perquè contenen els enzims responsables del
cicle de l'àcid glioxílic, una variant del cicle
de Krebs que permet convertir els greixos en hidrats de carboni.
Aquest procés només és present als vegetals
i alguns animals poc evolucionats; la resta és capaç
de transformar hidrats de carboni en greixos, però no pot
invertir el procés perquè no tenen glioxisomes.
Els glioxisomes són, doncs, orgànuls típicament
vegetals. El cicle de l'acid glioxílic és fonamental
per a les llavors de les plantes oleoginoses que acumulen reserves
en forma de greixos; durant la germinació es transformen
en glucosa.
5. Vàcuols
Els vàcuols de les cèl·lules
vegetals són compartiments separats del citosol per una
membrana. Generalment ocupen un 50% del contingut cel·lular
i poden arribar a ocupar fins un 95%. El conjunt dels vàcuols
d'una cèl·lula rep el nom de vacuoma.
Els vàcuols es formen en cèl·lules joves,
per fusió de vesícules derivades del RE i l'aparell
de Golgi. Estan relacionades estructuralment i funcionalment amb
els lisosomes.

Fig 12; Microfotografia d'una cèl·lula vegetal on apareix un gran vàcuol
que ocupa gran part del citoplasma
A les cèl·lules meristemàtiques lels vàcuols
són menuts i nombrosos, mentre que a les cèl·lules
més diferenciades, aquests es fusionen i ocupen la major
part del citoplasma. Entre les seues funcions destaquem:
- - Emmagatzematge de diferents tipus de substàncies:
productes de rebuig, substàncies de reserva, colorants,
verins, etc.
- Permeten que augmente la grandària de la cèl·lula per l'acumulació d'aigua: es tracta d'un mecanisme molt
menys costós qiue omplir la cèl·lula de citoplasma
- A les cèl·lules animals també poden aparéixer
vàcuols, tot i qie de menor grandària. Cal destacar-hi
els vàcuols pulsàtils, mecanisme que presenten
els protozous que viven en medis hipotònics per expulsar
l'excés d'aigua del seu interior.