НИИ медицинской генетики ТНЦ СО РАМН;
* Государственный научный центр ГосНИИ генетика, Москва;
** Экспертно-криминалистическое управление УВД Томской области
Изучен полиморфизм тандемного TCTA-повтора гена PAH у жителей Москвы и Западной Сибири. Выявлены 8 вариантов аллелей размером 228-256 п.н. Показано соответствие распределения частот встречаемости генотипов в обоих выборках равновесию Харди-Вайнберга и отсутствие статистически значимых межпопуляционных различий. Определены параметры информативности изученного локуса для использования в идентификационных исследованиях.
Ключевые слова: аллельный полиморфизм, микросателлитные повторы, ген PAH, полимеразная цепная реакция
В настоящее время в идентификационных исследованиях в практике криминалистики и судебно-медицинских исследованиях применяются молекулярно-генетические методы, основанные на генотипировании. Наиболее интенсивно изучаются полиморфные локусы генома, известные как тандемные повторы с изменяющимся числом копий, в частности микросателлитов или STR (shot tandem repeats) - с размером повторяющегося фрагмента 2-7 п.н. Такие локусы часто высокополиморфны, что и обусловливает их применение в медико-генетических целях. У американских европеоидов описан микросателлитный повтор в гене фенилаланингидроксилазы (PAH, хромосомная локализация - 12q22-q22.4 [3]), расположенный в 3-м интроне гена [3]. Данный STR-локус содержит различное число копий (повторов) последовательности 4 нуклеотидов - TCTA.
Задача настоящего исследования - изучить полиморфизм тандемного TCTA-повтора гена PAH у жителей двух различных регионов России (Москвы и Западной Сибири) и оценить параметры информативности локуса для идентификационных исследований в российских популяциях.
МЕТОДИКА ИССЛЕДОВАНИЯ
Работа выполнена на образцах ДНК, выделенных из крови неродственных представителей смешанной русскоязычной популяции Западной Сибири (Томск, Новосибирск) и популяции Москвы [4].
Методом ПЦР [7] исследовали область гена PAH, содержащую микросателлитные (TCTA)n-повторы. Олигонуклеотидные праймеры синтезировали фосфитамидным методом на ДНК-синтезаторе ASM-102U ("Biosset") согласно описанной структуре [3]: pr. F: 5'-GCC-AGA-ACA-ATA-CTG-GTT-C-3', pr. R: 5'-AAT-CAT-AAG-TGT-TCC-CAG-AC-3'. ПЦР проводили на амплификаторе "MiniCyclerTM" ("MJ Research Inc."). Реакционная смесь (50 мкл) содержала 0.1-0.2 мкг геномной ДНК, 40 пМ каждого праймера, 67 мМ трис-HCl, pH 8.8 при 200C, 16.6 мМ сульфата аммония, 4 мМ хлористого магния, 10 мМ 2-меркаптоэтанола, смесь 4 основных dNTP (по 0.2 мМ каждого) и 2 ед. Taq-полимеразы ("СибЭнзим"). После начальной денатурации (940C, 4 мин) 33 цикла ПЦР проводили в режиме: 930C - 40 с; 550C - 30 с; 720C - 1 мин. На последнем цикле стадию синтеза (при 720C) продлевали до 5 мин.
Продукты амплификации анализировали в 8% ПААГ длиной 20 см. На дорожку геля вносили 3-5 мкл реакционной смеси. Гели окрашивали бромистым этидием (0.5 мкг/мл). Для идентификации аллелей использовали соответствующие для данного локуса аллельные лэддеры и ДНК с известными генотипами. Визуализацию фрагментов ДНК осуществляли в УФ-свете с применением системы компьютерной видеосъёмки гелей на приборе "UV-VIS Imager II".
Частоты генотипов проверяли на соответствие равновесию Харди-Вайнберга по критериям c2 и G-статистики с помощью компьютерной программы RxC (Rows x Columns) на основе описанного алгоритма [5]. Ожидаемую гетерозиготность и параметры информативности изученного локуса: вероятность случайного совпадения генотипов двух неродственных индивидов (pM), среднюю вероятность исключения исследуемого образца по заданному генотипу (W), информационное содержание полиморфизма (PIC) рассчитывали с помощью компьютерной программы [1].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В исследовании генотипированы варианты микросателлитных TCTA-повторов гена PAH у 187 человек, представляющих 2 выборки из различных популяций России: русские жители Западной Сибири (n=115) и Москвы (n=72). Методом ПЦР в 3-м интроне гена PAH выявлены 8 вариантов аллелей (рис.1) длиной 228-256 п.н.
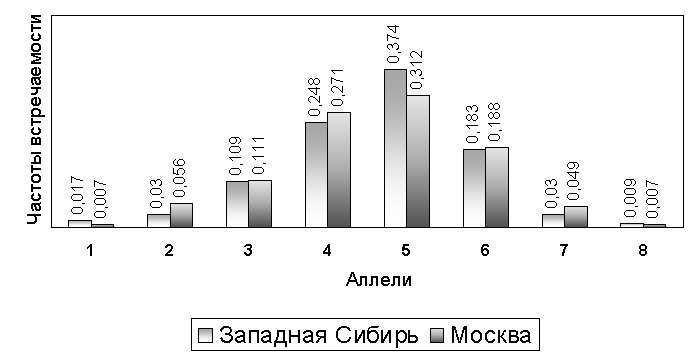
Полученное распределение частот встречаемости генотипов (табл. 1) соответствовало распределению Харди-Вайнберга в обоих популяционных группах (табл. 2). Это указывает на отсутствие внутренней гетерогенности изученных групп. Самыми распространенными были аллели 4-6. Спектр PAH-аллелей и их относительные частоты встречаемости были сходны в двух изученных выборках и в целом соответствовали описанному для европеоидного (американского) населения (рис. 2) [3].
Таблица 1. Частота встречаемости генотипов локуса PAH в двух популяционных выборках России - в этом файле не приводится
Рис. 1. Электрофореграмма анализа STR-полиморфных аллелей гена PAH в 8% ПААГ.
Дорожки 1, 9 - аллельные лэддеры; 2-8 - генотипирование неродственных людей.
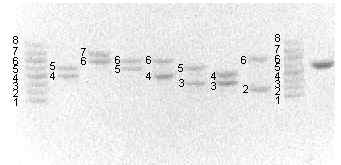 |
Рис. 2. Частоты встречаемости STR-аллелей в локусе PAH в двух популяционных выборках.
 |
Параметры аллельного полиморфизма свидетельствуют (табл. 2) о том, что изученный микросателлитный локус гена PAH является высокоинформативным для генетических исследований (молекулярная диагностика) и решения прикладных идентификационных задач. Генотипирование вариантов гена было апробировано в практике криминалистических экспертиз ЭКУ УВД Томской области, где показана их информативность при исследовании различных объектов (кровь, в том числе старые пятна крови и трупная кровь; слюна; окурки сигарет; кости, в том числе частично разложившиеся; образцы вагинального содержимого жертв) и возможность уверенного анализа аллелей как в препаратах высокомолекулярной ДНК, так и в образцах частично деградированной ДНК в количестве от 1 нг (иногда и меньше) до 50 нг, аллелей изученного полиморфного локуса гена PAH [2].
Таблица 2.Значения критериев полиморфизма, тестов c2 и G-статистики для локуса PAH в изученных популяционных выборках (M±m)
| Параметры | Западная Сибирь | Москва | Объединенные данные |
|---|---|---|---|
| Наблюдаемая гетерозиготность (Hobs) | 0,748 | 0,764 | 0,754 |
| Ожидаемая гетерозиготность (Hexp) | 0,751±0,040 | 0,776±0,049 | 0,762±0,031 |
| pM | 0,099 | 0,084 | 0,092 |
| W | 0,684 | 0,735 | 0,706 |
| PIC | 0,605 | 0,640 | 0,620 |
| c2 | 6,4651 | 10,1192 | 7,1238 |
| Вероятность | 1 | 0,9920±0,0028 | 1 |
| G-статистика | 8,0384 | 12,545 | 8,3582 |
| Вероятность | 1,0 | 0,9880±0,0034 | 1,0 |
Осуществленный нами сравнительный анализ с применением программы RxC в двух выборках не обнаружил значимых межпопуляционных различий частот выявленных генотипов (c2=21.0400 при p=0.7080±0.0144; G-статистика=25.4202 при p=0.7230±0.0142) и частотного распределения аллелей (c2=4.0871 при p=0.7980±0.0127; G-статистика=4.1048 при p=0.8100±0.0124).
Сходный характер аллельных профилей двух географически удаленных популяций России позволяет экстраполировать аллельные частоты, определенные в суммарной репрезентативной выборке (Москва и Западная Сибирь), на все русское население без риска существенно завысить потенциал индивидуализации. В табл. 3 приведены объединенные данные по частотам встречаемости аллелей локуса PAH, которые можно рекомендовать для использования в вероятностных расчетах при идентификационных исследованиях.
Таблица 3. Частоты встречаемости аллелей локуса PAH в русской популяции (M±m)
| Аллель | Длина аллеля, п.н. | Количество наблюдений | Частота встречаемости аллеля | "Потолочная" частота аллеля |
|---|---|---|---|---|
| 1 | 228 | 5 | 0,013±0,006 | 0,025 |
| 2 | 232 | 15 | 0,040±0,010 | 0,060 |
| 3 | 236 | 41 | 0,110±0,016 | 0,141 |
| 4 | 240 | 96 | 0,257±0,023 | 0,301 |
| 5 | 244 | 131 | 0,350±0,025 | 0,399 |
| 6 | 248 | 69 | 0,184±0,020 | 0,224 |
| 7 | 252 | 14 | 0,037±0,010 | 0,057 |
| 8 | 256 | 3 | 0,008±0,005 | 0,017 |
| Всего | 374 | 1,000±0,114 | 1,223 | |
Параметры аллельного полиморфизма свидетельствуют о том, что исследованный микросателлитный локус гена PAH у русского населения России является высокоинформативным для генетических исследований и решения прикладных идентификационных задач при идентификации личности, спорном отцовстве, криминалистических исследованиях и др.
ЛИТЕРАТУРА
- Ефремов И.А., Чистяков Д.А., Носиков В.В. // Молекул. биол. - 1996. - Т. 30. - С. 310-321.
- Одинокова О.Н., Кутмин А.И., Карташова О.Г., Пузырев В.П. // В кн.: Генетика человека и патология. - Томск, 1997. - С. 127-135.
- Goltsov A.A., Eisensmith R.C., Naughton E.R. et al. // Hum. Mol. - 1993. - Vol. 2. - P. 577-581.
- Johns M.B., Paulus-Thomas J.E. // Anal. Biochem. - 1989. - Vol. 180. - P. 276-278.
- Roff D. A., Bentzen P. // Mol. Biol. Evol. - 1989. - Vol. 6. - P.539-545.
- Saiki R.K.,Gelfand D.H., Stoffel S. et al. // Science. - 1988. - Vol. 239. - P. 487-491.