Transducción de señal - Muchos eventos intracelulares dependen de una perfecta coordinación intracelular y extracelular. La actividad metabólica y otros procesos biológicos intracelulares se encuentran bajo la influencia de señales químicas extracelulares como, por ejemplo, las hormonas.
La transducción de señal es un proceso por el cual se transmite un mensaje químico extracelular a través de la membrana plasmática para producir un cambio en la actividad metabólica dentro de la célula. El mecanismo varía de una hormona a otra, pero se puede trazar una serie de eventos generales en los que están implicados, al menos, tres tipos de proteínas:
-Proteína receptora - punto de unión con la hormona o alguna otra molécula efectora, de la que recibe el mensaje para la debida actividad metabólica.
-Proteína G - familia de biomoléculas que se unen a nucleotidos de guanina (GDP, GTP).
-Ciclasa de Adenilato - enzima que cataliza la formación de adenosina-3',5' -monofosfáto cíclico (cAMP), que es un segundo mensajero (molécula intracelular de corta vida que lleva la instrucción inicialmente transmitida por el primer mensajero, la hormona).
Resumen del proceso:
Una hormona o alguna otra molécula efectora se une con su proteína receptora situada en la membrana plasmática. El efector puede ser activante (+, Paso 1) o inhibidor (-, Paso 1').
Paso 2 - el receptor estimula la interacción con una proteína G, la cual activa la ciclasa de adenilato.
Paso 2' - un proceso inhibitorio inactiva la ciclasa de adenilato, vía interacción con una proteína G.
Paso 3 - la ciclasa de adenilato produce cAMP, que inicia una cascada de reacciones metabólicas, incluída la conversión de una proteína inactiva (posiblemente una enzima) en su forma activa.
Paso 3' - no se produce cAMP, por lo que no se produce la cascada de reacciones metabólicas.
En este proceso, la señal dada por la unión de la hormona al receptor se amplifica; es decir, su intensidad aumenta en cada uno de los distintos pasos. Sólo se requiere una molécula de la hormona que se une en la membrana para activar una molécula de ciclasa de adenilato; pero esta molécula enzimática puede catalizar la formación de muchas moléculas de cAMP.
Cada una de estas moléculas de segundo mensajero actúa para "desencadenar la activación" de las proteínas cinasa; así mismo, cada molécula de proteína cinasa activada actúa sobre muchas moléculas de la enzima blanco.
Debido a esta cascada de reacciones, la unión de una sola molécula de hormona conduce a una señal intracelular que puede amplificarse miles de veces. La señal aumenta aun más porque la célula blanco tiene muchos receptores para un tipo específico de hormona y cada interacción hormona -receptor lleva a la amplificación ya descrita.
GLUCOLISIS
Es el proceso más impórtante del metabolismo de carbohidrátos. De carácter anaeróbico, esta ruta la utilizan tanto los organismos anaeróbicos como los aeróbicos para generar una pequeña cantidad de energía en forma de ATP y NADH.
En los aeróbicos es la fase inicial para preparar a la glucosa en la producción de energía extra vía ciclo de ácido cítrico y fosforilación oxidativa. Esta última utiliza NADH como fuente de energía reductora para sintetizar ATP.
Las enzimas responsables de la glucólisis se encuentran en el citoplasma celular.
La vía glucolítica consta de diez (10) reacciones catalizadas por enzimas, usando Glucosa como sustrato inicial. Se divide en dos etapas principales:
Etapa I - primeras cinco reacciones; tienen como fin "activar" la glucosa. Es la etapa de inversión porque se consume energía (dos ATP).
Etapa II - ultimas 5 reacciones que comienzan con gilceraldehído 3- fosfáto y culminan con piruvato, ambos metabolitos de 3 carbonos. Es la etapa de "ganancia" porque se recupera la inversión inicial de dos ATP y se ganan otros dos.
A continuación se ilustran la reacciones de la vía glucolítica:


Metabolismo de Piruvato
El factor principal que determina el destino final del piruvato es la disponibilidad de oxígeno:
(a) En las células aeróbicas con abundancia de oxígeno, el piruvato sigue una vía de degradación oxidativa a través del ciclo de ácido cítrico y de una fosforilación oxidativa hasta que cada carbono se oxida a CO2 y toda la energía potencial se captura en forma de ATP. Los electrones separados de los sustratos oxidados los capturan las moléculas de NADH y FADH2 y los transfieren al último aceptor de electrones: oxígeno. Los dos moles de NADH producidos en la glucólisis en el paso de gliceraldehído-3-fosfáto a 1,3-difosfoglicerato se usarian para esos efectos.
(b) En ausencia de oxígeno (anaeróbico), se produce la fermentación láctica:

Este proceso permite el reciclaje del par NAD+/NADH + H en sistema anaeróbicos.
Entrada de otros Carbohidrátos a Glucólisis:


2- Hígado - como variedad de almacenamiento de glucosa, sufre una ruptura fosforilítica catalizada por la fosforilasa de glucógeno (glucogenolisis). Esta reacción es más favorable que la anterior.

La gucógeno fosforilaza va rompiendo progresivamente los enlaces α(1→ 4') de la cadena de glucosas del glucógeno, liberando moléculas de glucosa-1-fosfáto. Esta última se convierte en glucosa-6-fosfáto por una fosfoglucomutasa.
En el hígado, la glucosa-6-fosfáto se convierete en glucosa por la acción de la glucosa-6-fosfatasa. La glucosa se mobiliza a la sangre y de ahí a los tejidos que la requieran.

La gluconeogénesis (síntesis de glucosa a partir de precursores que no son carbohidrátos) se lleva a cabo en el hígado y en la corteza renal.
La glucosa se sintetiza principalmente en el hígado y se transporta hacia el músculo a través de la sangre. El músculo esquelético degrada glucosa por la vía de glucólisis y fermentación láctica. Después el lactato se transporta al hígado para usarse en la gluconeogénesis.

El uso primario de la glucosa sintetizada por gluconeogénesis es el del catabolismo por tejidos nerviosos y la utilización por el músculo esqueletal. Glucosa es tambien la materia prima para todos los otros carbohidrátos, incluyendo las amino-azúcares, los polisacáridos complejos y los componentes carbohidrátos en glucoproteínas y glucolípidos.
Tejidos que usan glucosa como única fuente de energía (carecen de las enzimas necesarias para la gluconeogéneis):
GLUCOLISIS (→) VS GLUCONEOGENESIS (←)

Piruvato y Lactato en Gluconeogénesis:
(a) Utilización de piruvato del citoplasma para sintetizar fosfoenolpiruvato con enzimas del citoplasma y de la mitocondria.

(b) utilización de lactato del citoplasma para sintetizar fosfoenolpiruvato con enzimas del citoplasma y de la mitocondria.

Se requiere también un señuelo (primer) en la forma de un oligosacárido de α(1→ 4') glucosa unida a una proteína (glicogenina). El grupo glucosilo de UDP-glucosa se añade a la cadena polisacárida por la enzima glucógeno sintasa:

La regulación de las rutas metabólicas es vital para mantener constantes los niveles de ATP: el ATP debe formarse tan pronto como se utiliza. La manera más efectiva de lograrlo es controlando los niveles de relativos de glucosa libre y almacenada.
Si en determinado momento la célula necesita energía, entonces la glucólisis debe entrar en acción y degradar glucógeno, liberando glucosa-1-fosfáto. Si por otro lado no se necesita energía, la vía glucolítica debe cerrarse y almacenar la glucosa para utilizarse después.
La regulación metabólica es posible gracias a la regulación enzimática vía:
Alta concentración de ADP y AMP indica un bajo nivel de energia induce a la glucólisis.
Los sustratos y productos de las reacciones también funcionan como efectores.
PUNTOS DE REGULACION
(I) Glucógeno sintasa y glucógeno fosforilasa:
Estas dos son parte de una enzima bifuncional que se encuentran particularmente en el hígado y músculo esquelético. En ambos lugares se regulan de modo distinto porque el glucógeno, como sustrato juega un papel distinto en cada tejido: en el hígado se almacena el glucógeno para todo el organismo: cuando se necesita glucosa, la fosforilaza (glucogenolisis), movilizándola hacia la sangre para ser distribuída en todos los tejidos; cuando hay exceso de glucosa, la sintasa se encarga de la síntesis de glucógeno a nivel hepático(glucogénesis).
En el músculo, los bajos niveles de glucógeno se degradan por ruptura fosforilítica(glucogenolisis) y glucólisis para obtener ATP local para la contracción muscular.
La glucógeno sintasa y la glucógeno fosforilaza son proteínas diméricas y existen en dos formas intercambiables: forma activa e inactiva. La interconversión de las dos formas enzimáticas de ambas enzimas está controlada por las hormonas adrenalina (epinefrina), glucagón e insulina.
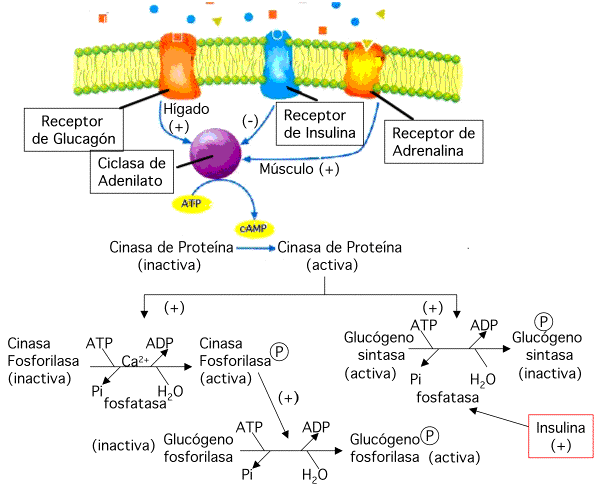

Efectos hormonales:
Glucagón - actúa en el hígado; estimula la glucogenólisis y gluconeogénesis; desactiva la glucogénesis.
Adrenalina - actúa en los músculos y estimula la glucogenólisis y glucolisis, y desactiva la glucogénesis. En el hígado estimula la glucogenólisis y gluconeogénesis; desactiva la glucogénesis.
Insulina - actúa en el hígado, los músculos y el tejido adiposo inhibiendo la degradación de glucógeno (glucogenólisis) y promoviendo la glucogénesis en hígado y músculo (disminuye niveles de glucosa en la sangre).
Resumen:

Esta enzima regula la entrada de glucosa en la célula. En el músculo, esta enzima se inhíbe por su producto, Glucosa-6-P, por medio de la inhibición por retroalimentación: si se acumula mucha glucosa-6-P, el flujo glucolítico disminuye y se conserva la glucosa.
Como en el músculo no hay glucosa -6-fosfatasa (que convierte glucosa-6-P en glucosa), la glucosa-6-fosfáto se ve obligada a a seguir la ruta glucolítica para la formación de ATP. Esta enzima tiene una alta afinidad por glucosa (km = 0.1 mmol/L), por lo que fosforilará rápidamente la glucosa. Esto asegura que aun con una baja concentración de glucosa en el músculo (0.1 mmol/Lt), la reacción de la hexocinasa puede proceder a la mitad de su máxima rapidez .

En el hígado (y páncreas) hay otra isoenzima, la glucocinasa. La glucocinasa tiene un km de 10.0 mmol/L, por lo que puede copar con una alta concentración de glucosa; así ayuda al hígado a mantener los niveles de glucosa en la sangre.
Contrario a la hexocinasa, la glucocinasa no es inhibida por glucosa-6-P; esto asegura que cuando el hígado recibe una gran cantidad de glucosa, ésta se puede fosfolirar a glucosa-6-P previo a la glucogénesis.
Fructosa-2,6-bifosfáto, que se produce por la acción enzimática de la PFK-2, es el regulador más importante de glucólisis.
Fructosa-2,6-bifosfáto es un activador de la PFK-1, la enzima que cataliza la formación de Fructosa-1,6-bifosfáato a partir de Fructosa-6-fosfáto, paso que favorece la ruta glucolítica.
PFK-2 (fosfofructocinasa-2) y FBPasa-2 (fructosa-2.6-bifosfatasa son parte de una enzima bifuncional. Sin fosforilar, la PFK-2 está activa y la FBPasa-2, inactiva.
(A) Hígado:
La actividad de PFK-2 está unida a la acción de glucagón vía cAMP (respuesta a bajos niveles de glucosa), que causa la fosforilación de PFK-2, inactivándola y la activación de la FBPasa-2.
Esto lleva a la degradación de Fructosa-2,6-biP a Fructosa-6-P lo que causa una disminución de la concentración de Fructosa-2,6-biP, la inactivación de PFK-1, y se elimina la inhibició de la fructosa-1,6-bifosfatasa (FBPasa-1).
Como consecuencia se reduce la glucólisis y se activa la gluconeogénesis. (La acción de glucagón vía cAMP también causa la activación de la fosforilasa de glucógeno, lo que promueve la glucogenolisis, aumentando la concentración de glucosa).
PFK-1 = ATP es sustrato e inhibidor alostérico
PFK2 = se activa por Pi y se inhibe por citrato

(B) Músculo:
La PFK-2 activa la glucólisis porque aumenta la concentración de Fructosa-2,6-bifosfáto, que es el regulador más importante por ser un activador de la PFK-1.
La actividad de PFK-2 está unida a la acción de adrenalina vía cAMP (respuesta a la necesidad de energia), que causa la fosforilación de PFK-2, activándola y la inactivación de la FBPasa-2.
Esto lleva a la formación de Fructosa-2,6-biP a partir de Fructosa-6P, lo que causa un aumento de la concentración de Fructosa-2,6-biP, y la activación de PFK-1.
Como consecuencia se aumenta la glucólisis. (La acción de adrenalina vía cAMP también causa la activación de la fosforilasa de glucógeno, lo que promueve la glucogenolisis).
El músculo no es un tejido gluconeogénico por carecer de la enzima glucosa-6-fosfatasa. que cataliza la reacción de glucosa-6-fosfáto a glucosa.

La cinasa de piruvato del hígado se inhibe con altos niveles de ATP y, por tanto, hace más lenta la formación de piruvato. La cAMP (vía glucagón) convierte la cinasa en su forma inactiva. De esta forma se inhibe la glucólisis y se activa la gluconeogénesis hepática.

