Ancillary Articles
for the New Model of EvolutionShort articles explaining stepped and large-scale changes, primitive life, behavior first, use of evolutionary terms and complex time.
 (Back to Top.) ____________________ (Site Navigation Page.)
(Back to Top.) ____________________ (Site Navigation Page.)![]()
Return to:
1.0 The Theory of Large Changes 2.0 The Theory of Complex Accumulations
3.0 The "Peripheral Niche" Mechanism 4.0 The Theory of Phylogenic Evolution
5.0 The "Smart Gene" Hypothesis 6.0 A 'Two-Axis' Model of Evolution
Introduction Contact Information
![]()
Introduction
Living nature evolves by a process of variation, adaptation, fitness, selection and reproduction. But humans, the inquisitive creatures, impose on nature the additional requirement that this process of evolution makes logical sense. The process of small-scale evolution has been verified many times in experiments and mathematical modeling to make impressive logical sense. From this triumph humans have imposed an additional requirement that the process by which small-scale changes work must also apply for the large-scale changes. But many large-scale effects, like evolution of new novelties such as sex, the origin of new species and phyla, or the evolution of human behavior cannot be explained this way. It seems that some process is interfering with the logic of transposing small-scale evolutionary changes into large ones.
The new theory proposes that the difficulty might be a simple one. (Note: Just the explanation of why we cannot linearly transpose small to large-scale changes is a simple one. Evolution itself is still a complex process.)
So, small-scale changes are not changes over short times or genetic distances times, but adaptations of easy-to-modify traits. But once we transpose adaptation over large time scales and genetic distances we include evolution of hard-to-alter attributes, or traits once easy-to-alter now matured into hard-to-alter ones. Only this transposition from small to large is not linear in time or genetic distance. Lungfish barely alter over huge times, but human morphology and behavior changed markedly in a short time over a short genetic distance. So comparing how species alter or traits evolve by measuring changes over time does not reveal the scale of the change. Instead, we need to measure the relative difficulty of evolving traits, regardless of time.
We can measure the difficulty evolving traits by an effect of fitness. In one sense, all DNA-coded traits are easy-to-alter, and all successful adaptations are fitness gains. But some easy-to-alter small changes are hard to accumulate over long distances, and result in an eventual fitness loss to descendents from the original fitness held by ancestors. (Critics object that they cannot "see" how small fitness gains could result in a fitness loss. There is evidence that fitness is less for complex organisms, which evolved later, than simple ones, which evolved first. So organisms can evolve against a fitness loss as a fact. The difficulty is explaining why.) Large changes in evolution then, are not just of scale. They are small accumulations over large distances that result in a fitness loss to host lines. These changes are distinct events in the history of life, which impose a pattern on all large-scale evolution. (This is a phylogenic explanation. An asteroid strike also causes large-scale changes, but not because of fitness, or whether traits are easy or hard to evolve. See use of terms.)
The new theory is broken into the following sections:
1.0 The Theory of Large Changes explains that any evolving system contains units with both easy and hard to change attributes, and this applies to living evolution. Although attributes differentiate into easy or hard to evolve traits naturally, it will still make evolution more efficient. This is because fitness allows the easy-to-adapt traits to radiate as a huge variety, but will restrict evolution of new, hard-to-evolve traits to singular, concentrated efforts.
2.0 The Theory of Complex Accumulations explains the arguments and evidence supporting the view that fitness will fall as organisms evolve greater complexity. Organisms can still evolve against a fitness loss by accumulating many low yield fitness gains, not normally profitable under stable competition. The final part will explain why this loss of fitness for complex accumulations will produce the stepped effect of evolutionary change.
3.0 The "Peripheral Niche" Mechanism explains that most isolating mechanisms used in conventional theory would already be strong enough to force evolution against shallow fitness losses. However, evolution (in the past) of major new phyla and classes must have occurred against huge fitness losses which known mechanisms could force. Yet also in the past there would have been environmental ranges that had to be penetrated by each major taxonomic group a first time. Organisms penetrating these new ranges the first time could encounter very steep fitness declines, which might account for how major new novelties could evolve against steep fitness losses.
4.0 The Theory of Phylogenic Evolution argues that species could only be forced into fitness declines by some form of evolutionary saturation amongst existing types, but this has never been explained. Only saturation cannot occur among species, because species are collections of individuals, and individuals contain continuously variable traits. However, we can redefine a phylogeny as an exclusive set of non (or slowly) varying traits. We can then prove that unlike for species, phylogenies would evolve towards saturation, beyond which they cannot evolve further.
5.0 The "Smart Gene" Hypothesis deals with the problem that if all DNA-coded traits are easy to alter, why would the effect of some attributes being easy but others hard to change persist in DNA-coded life? The answer is that even from a "selfish gene" perspective, genes expressing proven, homologous traits will not want to alter sequence. They will instead obtain greater propagation by resisting change, and radiating throughout a huge variety of types.
6.0 The 'Two-Axis' Model of Evolution teaches that the contrast between the small and large-scale changes is easier to explain on a 'two-axis' model of evolution. The second axis is to demonstrate that while over short distances homologous traits appear "fixed", over very large time-scales "fixed" traits are evolving too, in much slower cycle times.
In addition;
Large Scale Changes explains the difference between large-scale and small-scale changes in evolution.
Use of Terms explains why some terms used might appear different from the usual meanings, and welcomes suggestions for clarifying or improving the use of terms.
Primitive Life provides some background on the impetus of evolution, as explained by the new theory.
Stepped Changes shows a diagram of "stepped" evolution of mammals, birds and dinosaurs, and some of the difficulties trying to distinguish the actual "stepped" changes.
Complex Time suggest a further, interesting direction where the above arguments might lead, though not pursued here.
![]()
Complexity evolves in steps, once all the simple adaptations of easy-to-alter attributes at an existing level of complexity are consumed. Only this is disputatious, because it oversimplifies the evolutionary pattern of complex life. Mammals say, even if we allow that they are more complex than dinosaurs, still did not evolve from them! On the contrary, reptilian ancestors of mammals first evolved in parallel, or slightly before the ancestor of dinosaurs (see diagram). Mammal complexity also accumulated slowly, spanning the entire age of dinosaurs.
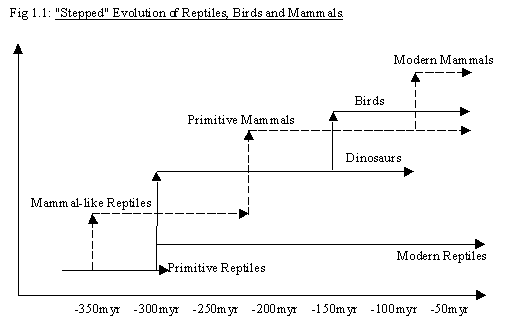
However, while the pattern of life seems to dispute the theory, this is life's modern complexity. The pattern is still there. In primitive life it seems certain that prokaryote life preceded more complex eukaryote life, perhaps by billions of years (some people dispute this) or single-celled preceded multi-celled organisms, and asexual preceded sexual reproduction. More of a problem is that initial designs are only potentially more complex. There seems to be nothing to indicate a Synapsida (ancestor of mammals) is a more complex design than a Diapsida (ancestor of dinosaurs). If anything, they seem to have evolved about the same time.
A related difficulty is the radiation pattern of the famed Cambrian 'explosion'. Steven Gould and others have asserted that most body plans evolved at one time during the Cambrian. After that, evolution became more a pruning of unsuccessful designs than the familiar branching tree of life. This is partly possible. (My favorite analogy is the early days of airplanes. There were hundreds of weird early designs, but only a few ended-up true generic ancestors of modern airplanes.) However, there were still many branching radiations at sub-phyla level after the Cambrian, as indicated in the diagram. Plus the new theory offers other reasons why new phyla would not emerge post Cambrian. (See Theory of Phylogenic Evolution.)
(Note: Above diagram is also grossly simplified. Any corrections, suggestions, or criticism this diagram, please let me know. See Contact Information.)
![]()
In current theory the differences between small and large-scale changes in evolution are ones of scale.* If a small change accumulates over a long time, it produces an effective large change. This is easily understood. (*Note: Again, this is the phylogenic difference. Asteroid strikes, continental drift, climate change, migration, are all large-scale changes that also effect how phylogenies evolve, but do not occur for phylogenic reasons. See Use of Terms.)
Only in evolution some attributes are easier change than others. It is easier to vary the shape of a wing, which occurs all the time, than evolve the material "feathers", which only evolved once. It is easier to adapt the function "limbs" into arms, legs, flippers, and wings, which occurred many times, than evolve the four-limb body plan, which only occurred once. So, small changes over a long time will transform a flipper into a leg, or an arm, a wing or even back into a flipper again. This process is not in dispute. But once it has evolved the first time, no amount of small changes will allow a four-limb body plan to evolve again, nor will small changes allow feathers to evolve again. And to understand the large-scale processes of evolution, we need to understand why this is.
Diagram shows how large-scale changes appear typically in 'two-axis' model.

![]()
Early expositions of this theory were harshly criticized for inappropriate or confusing use of terms. I have tried to correct this, but many terms from existing theory do not express quite the idea intended.
The biggest difficulty is terms to distinguish the "easy-to-evolve" from "hard-to-evolve" traits. I now use homoplastic to mean easy-to-evolve, but previously I used 'analogous', which was sharply criticized. (I believe I was correct. Analogous traits are easy-to-evolve, which is why they reappear in many forms.) For "hard-to-evolve" I have used homologous throughout. (Strictly, hard-to-evolve traits first mature as traits once homoplastic, and do not become homologous until they radiate into many lines.) Terms I prefer are polyphyletic for easy-to-evolve, and monophyletic for hard to evolve, because these convey an "easily repeatable" and "single origin" meaning. Again, I have been sharply reminded that these terms apply to taxonomic classifications, not to how traits evolve. I welcome suggestions of better terms to use.
More serious, in early writings I used macroevolution to describe evolution of the hard-to-evolve monophyletic traits and microevolution as evolution of the easy-to-adapt, polyphyletic traits. Again, I believe I was correct, because large scale changes in life ultimately result from evolution of fresh monophyletic novelties, which produce a radiation of types by easy-to-adapt changes of the basic plan. However, this definition overlooks that large-scale changes also have non-phylogenic causes, such as geological change. (Change of climate, geology, and astronomical impacts now seem to be the focus of macroevolution.) So, the process I previously described as macroevolution I now refer to as phylogenic or homologous evolution. This is a contributing, but never the sole cause of above species level change.
Only many critics hold that regardless of terms, differences between micro and macroevolution are a human classification of scale, but not different processes of nature. Here we do disagree. Regardless of what we call it the evolution of hard-to-evolve, homologous traits is an episodic, monophyletic process involving an eventual loss of genome fitness over the lineage hosting the change. The process is also progressive, and evolves towards saturation of its possibilities. This is not the way non-progressive, polyphyletic, easy-to-alter change occurs. Only in advanced types so-called homologous traits do evolve from traits once homoplastic, by processes that appear identical to microevolution. This is why we need a 'two-axis' model. The type of change we identify as special or different (let us call it macroevolution) involves a "phase shift" in the cycle times of the evolution of traits, and current models of evolution do not identify this type of change.
Most other terms used are defined in the relevant sections. If I did misuse an established term, please accept that it might be ignorance of usage, but no obfuscation was intended. Any help, corrections, or suggestions in sorting out these terms is always most welcome.
![]()
The theory of "Behavior First" is an explanation of why flexible behavior can lead to faster adaptation of genotype. The theory is not Lamarckian, but it does remind people that early giraffes were not motivationally forbidden to stretch their necks if they wanted too. Only stretching necks does not alter genotypes. Rather, early giraffes phenotype adapted by stretching necks "first", but later Darwinian selection weeded out individuals not as well genotype adapted as they are phenotype adapted. (Randomly born long-necked giraffes procured more offspring, and genotype adapted.) Similarly, we say that mammals can adapt faster than reptiles, but not because their genes mutate faster. Mammals alter behavior faster, so they adapt behavior "first". Altered behavior does not change genomes, but it presents Darwinian selection a broader variety from which to choose the fittest, which does alter genomes. (Of course many genes in mammals evolved later than genes in reptiles. Some of these might not so much mutate faster, but find ways to express greater evolvability, or they can change when change is required.)
![]()
Until more is known, all theories of primitive life must be speculative. But by whatever way primitive life evolved it must reflect two tendencies, fundamental not only to living evolution, but properties of the universe. The first is a tendency for some properties of matter, life, geometry and existence to be stable but hard to change, and others to be less stable but easier to alter. (See Theory of Large Changes or 'Two-Axis' Model for examples.) The next tendency is controversial, as it has never been quantified. This is a tendency of information to 'replicate'. We know information replicates for systems that are already alive. And we know that non-living information from the early universe on the polarity, charge, density, and structure of matter has been replicated throughout existence. But no physical law of how replication of non-living information transforms into living self-replicating information has been formulated.
We can suppose that the unifying law (not yet discovered) concerns transformation of energy into structure. Roughly, the early universe contains superabundant energy. Part of this energy is dissipated from its initial concentration by conversion (condensation) into stable, enduring structures. (The condensed energy will later to be reconverted back into energy aeons hence, when the energy stocks of the current universe decline.) There is inferential evidence of this (which we will not go into) from cosmology and microphysics, but the process makes sense in evolution. Roughly, energy can only be absorbed into structure up to a certain level of complexity, such as amino acids and other building blocks of life. Beyond that, the Second Law will degrade structures as fast as they can form, so structures need to self-replicate to be maintained. (This might explain why early cells evolved in high pressure and temperature, where energy was abundant.) And while this still does not explain how life first evolved, it could help explain how later life evolved. The issue is that early life was already good at replicating copy by being structurally simple. But after inception life continued to evolve into increasingly complex forms. Only complex life is harder to replicate, and harder to replicate enduring copy at the DNA level. (We are not sure if DNA was the initial impetus. It was more likely RNA or some polymer combinations, because ribosomal RNA and basic proteins seem more widely replicated than many DNA sequences.)
Homologous traits begin life (literally) then not as "genes", or even DNA, but as a pre-biotic property of molecules that some structures were more stable and enduring than others. Not until life evolved to a sophisticated state of coding its efficiency in double helix DNA (an evolved, advanced code) did genes emerge, though we are not sure when. Chemically, any sequence of DNA seems as good as any other. (This might not be correct. DNA is more stable as a three-letter than two-letter code. DNA also folds into a three-dimensional structure with some shapes more stable than others, and so on.) But DNA only evolved as part of cells and cell replication machinery. The cells, sugars, proteins and RNA existed first, and DNA was a selected for a need. (There are theories of "naked" DNA first, but this is unlikely for many reasons.) Moreover, there is a minimum of DNA necessary to code the simplest cell that can exist. Only that cell must be very fit, because it must replicate exactly. (There probably never was a perfectly designed minimum complexity cell. Only if such an entity could exist, to retain minimal complexity it must replicate exactly. See argument in Fitness Does Fall.) (See also "Smart Gene" Hypothesis.)
In early life then "smart" genes performed essential roles in larger, more complex cells.
Note on gene selfishness. Materials in contact for long periods develop 'stiction'. But DNA materials are constantly renewed and only the information content is conserved, so 'stiction' would not apply. (And it certainly would not apply to radiating sequences!) More likely, the duplicated and edited code in eukaryote DNA keeps a 'history' of changes and knows which sequences to conserve. And if the efficiencies seem small they did evolve in early life. Here long periods were available to accumulate small gains but minuscule gains could yield huge fitness rewards in subsequent evolution. (If a 0.01% gain of fitness allowed a type to become the founding design of all subsequent life, the effort would be worth it.) Maintaining a sequence because it was already stable allows renegade "selfish" genes to exploit the attribute, but early competition would find an optimum. If anything, modern genomes show a balance weighed towards replicating more sequences than required for expression.
![]()
Several authors have independently raised the issue of complex time, though not directly related to living evolution. Steven Hawking has mentioned it as a theory of the universe. Frijof Capra in his latest "Web of Life" mentions complex numbers, but more in relation to fractal theory. For myself, who began with the study of human evolution, it seems that the change in human evolution from one of adapting biology to one of adapting culture was a "phase shift". It would also seem intuitively from this phase shift that humans continue biological evolution in complex, rather than real time. The fact that humans were evolving biologically in complex time could also help explain many human obsessions, such as a search for immortality, or a spiritual life beyond this one (although there are other ways to explain this).
The 'two-axis' model of evolution raises the possibility of introducing complex time directly into evolution, although its physical interpretation is in this case mundane. Complex (or 'unreal' time) here just means that the passage of time is no longer affecting certain attributes, and this is the case. Evolution of the DNA code or basic cell structure is complete for present life on Earth. The passage of time does not affect how these alter further. In the 'two-axis' model we show any attributes unaffected by time as "rotated" 900 away from the 'axis' of time. (Time is at 00, or flowing left to right across the page, which is human convention.) However, we could as easily describe rotation out of the plane of time as evolving in 'j' (or Ö -1) time. The mathematical conventions 900 and Ö -1 are here interchangeable. Only we have a less mystical idea of what the 900 means, so this is the convention used in the body of the text.
The other possibility for introducing complex time concerns the evolution of complex or non-linear systems. The new theory contests that because some attributes are easy, but others hard to alter, this will induce a stepped effect in the evolution of any system. Basically, it will be more efficient for evolving systems to consume the easy-to-enact changes first, and only evolve hard-to-alter traits in concentrated efforts. This author believes this could be proven for a system evolving "a + jb" elements, rather than just "a" elements, but lacks the skills to prove it. (I will pursue this. Any experts out there in complex mathematics who would like to comment?)
At this stage this author is unsure if anything will come from introducing complex (Ö -1) time into evolutionary theory or human behavior. But the challenge is an interesting one.
![]()
Return to
New Model of Evolution Home Page