Palindromati
Fernando Castro-Chavez.
fdocc at yahoo dot com
Independent
Biotechnologist.
Click here for the Spanish Version (en Castellano).
Reciente Entrevista (en español).
Recent interview (English.)
ABSTRACT
This article describes a family of artificial
heterotranscripts (RNA chimaeras) composed by thousands of Genbank sequences containing fragments or the complete EcoRI-like adapter acting as the
palindrome linker ctcgtgccgaattcggcacgag,
binding together two or more genes that may be produced by different
chromosomes. This happens due to current methodologies producing the reported
sequences, found in the Genbank, in Affymetrix microarrays, and in many
published articles reporting or using those sequences that include the EcoRI-like linker inside coding regions,
and/or 5'UTR or 3'UTRs mRNA sites. This EcoRI-like linker and its heterotranscripts are
here deemed as experimental artifacts, characterization that can be helpful to
prevent errors, both in the studies of molecular mechanisms and in the drug
discovery process.
Key words: EcoRI, Palindrome,
Perilipin, Genbank, Affymetrix, microarray, species-specific.
INTRODUCTION
It is
vital in the discovery of new medical treatments to target precise molecules
without having side effects for organic tissues. To accomplish this objective
it is necessary a stringent quality control within molecular databases. This
article describes the finding of numerous methodological artifacts reported to
the Genbank. It is recommended a most
carefully analysis of nucleic acid sequences for biological, medical and drug
discovery purposes.
A single RNA binding in
one-strand two different genes from two different human chromosomes (1) was the
theoretical beginning for the study on heterotranscripts.
Here, I
define heterotranscripts as
chimaeras, sequences composed by fragments corresponding to two or more genes
from the same or from different chromosomes.
I thought that such a
phenomenon reported in reference (1) must have been reflected in a rational and
logical combination of Intelligently Designed gene products (2, 3). As most of
the vital molecules and biological pathways are present in many organisms, I
initially thought that the phenomenon described in reference (1) maybe should
be present in many natural sequences as a possible functional common
denominator.
I initially supposed that the
study of sequences similar or related to the one present in reference (1) maybe
could be helpful for our understanding of the molecular basis to biological
change.
Thus, this particular
phenomenon was a possible prospect for the abundance of proteins exceeding
their number of genes via multiple modular combinations of diverse mRNAs.
Recent estimates for humans reckon above a million of proteins produced by just
20,000 to 25,000 genes (4).
With these considerations in
mind, my initial idea was that, if reference (1) was true, then the production
of those numerous proteins could have had a putative process of RNA
hetero-linkage at their formation.
However, after five years of
comparing sequences, I came to realize that those hetero-sequences using that
same oligonucleotide as its common linker were just methodological artifacts.
The
common element of these chimeras is the linker ccgaattcgg (as presented in reference 1 inside the
sequence L21934 for the H. sapiens ACAT-1
enzyme). This leaves references 1, 5 and 6 (if real), as a one and unique
possible species-specific phenomenon
in humans (2, 3).
Another
article on the same sequence (1), has recently been published by the same group
(5). Its authors mentioned since reference (1) the similarity of that linker
with the EcoRI-adapter (a tool
extensively used in molecular biology research), so the door still is open to
verify whether this is a methodological artifact or not (5).
The
initial construction of that sequence demonstrates that their cDNA library was
transformed in E. coli (strain MC1061) using the phagemid vector pBluescript as well as with the
expression vector pcDNA. Then, they retransformed
it in the same E. coli strain (6), again.
However, I have recently seen that the use of similar vectors can be
involved in the production of chimerical artifacts in multiple instances, like
in those examples presented in Tables 1 and 2 (7).
A
possible, however remote, explanation for reference (1) is that we are dealing
with a natural process, mostly restricted to humans. Yet, whatever the final
verdict may be, the fact is that the EcoRI-like
linker or adapter described in (1) was the starting point for the next
findings, described in this article.
My
personal hypothesis is that heterotranscripts or chimeras including the EcoRI-related palindromic linker ctcgtgccgaattcggcacgag or its related sequences, extending
themselves to at least twelve bases, are artifacts from the molecular
methodologies used, mainly mediated by its host-vector interactions.
RESULTS AND DISCUSSION
The finding of a related palindrome in
Affymetrix microarrays
The
basis for this article appeared while working with antiobesity microarrays. By
studying the changes of gene expression in the obesity resistant perilipin
knock out mice (8, 11), with the DNA-Chip Affymetrix
MG-U74A-v2, analyzed using the free educational software dChip V.1.2 (9). One particularly intriguing hetero-transcript was
the nucleotide sequence AB030505, initially reported by its submitters as the Mus musculus mRNA for UBE-1c1, UBE-1c2
& UBE-1c3 (complete cds). The following paragraph describes the sequence
AB030505 and the common EcoRI-like
linking element present in thousands of other Genbank sequences.
A
careful study of the nucleotide sequence AB030505 using Blast (10) led me to an element that was linking two large sections
from two different genes:
1. The nucleotide sequence AK078792 from chromosome 10,
coding for a melanoma ubiquitous mutated protein homologue (Mum1) and
2. The nucleotide sequence BC036273 from chromosome 12,
coding for retinol dehydrogenase 11 (similar to Arsdr1). The linking element
within the sequence AB030505 corresponded to the palindrome ctcgtgccgaattcggcacgag,
composed by 22 nucleotides.
Here
again, as in the initial report (1), two transcripts originated in two
different chromosomes were linked together in one mRNA strand. Those 22 bases
contain the core palindromic linker ccgaattcgg at its center, which is
similar to the one initially reported by reference (1).
A
palindrome sequence for the double helix of DNA has the same nucleotides if
read from 5' to 3', which is the normal reading direction, either from the plus
(+) or from the minus (-) strand. A manual and visual assessment of this
palindromic linker was done. Amazingly, this linker was present in thousands of
sequences reported to the Genbank.
In the
full Table 2 (7), I present many examples of the palindrome (or related
sequences) being reported as if they were present inside coding regions. The
palindromic linker mentioned is frequently translated as the artificial peptide
RAEFGT, absent in sequenced protein
databases (10).
Increase in the number of palindromic sequences reported to
the Genbank
A monthly increase was seen in
the number of sequences containing the EcoRI-like
linker or its derivatives inside thousands of sequences. In one recent example
(14 Oct. 2005) done in Blastn (nucleotide to nucleotide alignments),
selecting the non redundant (nr) nucleic acid database sequences of Genbank, a query of 44 palindrome
letters was used:
CTCGTGCCGAATTCGGCACGAGCTCGTGCCGAATTCGGCACGAG
With
this query, I obtainined 6010 Blast Hits using the next query conditions:
1. 106 as the minimum expected number. Some
results are presented in Table 2 (7).
2.
1000 as the number of descriptions and of alignments.
In the Genbank's alternate database containing expressed sequence tags (est),
which are mRNAs for putative proteins, the number of sequences containing the
palindromic EcoRI-adapter is also
present by the thousands.
Additional palindromes found by using microarrays
Additional targets pertaining to
these linkers were also found while studying the results of microarrays
available online using the software
tool dChip (9) coupled to the Affymetrix probes databases. Table 1
shows examples containing the palindromic linkers.
Affymetrix has been a
successful microarray methodology, i.e.,
to evaluate the gene expression in humans, mice, and rats. However, both the
presence of artificial heterotranscripts and/or of their own artificial linkers
can lead to a misrepresentation of its real expression inside the tissues, as
the area under the curve is reduced for those genetic sequences.
Table 1. The EcoRI-related palindromic linker is
present both in the Genbank sequence
targets and in Affymetrix microarray probes for humans, mice and rats.
|
ID/ Organism
|
EcoRI Affymetrix Probes
from Genbank [DNA Chip]
|
Graphic, non-expression of EcoRI-linker
|
Reference
|
|
AB002533_at
Homo sapiens
|
gaattcggcacgagcacgcgtgaga,
ggcacgagcacgcgtgagacttctc
[in the Human DNA
Chips HuGene-FL and in Hu6800]
|

Ribosomal protein LP2 (Qip1)
|
Shipp et al, Nature Med.
8, 68. 2002
|
|
AJ243503
(99534_at)
Mus musculus
|
ttcggcacgagctcgtgccggtcct
[in the Mouse DNA
Chips MG-U74Av2 and in MG-U74A]
|

Adipocyte
Ghrelin
|
Castro-Chavez et al. Diabetes 52, 2666. 2003
|
|
AI045710
(rc_AI045710_at)
Rattus norvegicus
|
atgatatgtacagatccctcgtgcc,
tgatatgtacagatccctcgtgccg,
tatgtacagatccctcgtgccgcct,
tgtacagatccctcgtgccgcctcg,
gtacagatccctcgtgccgcctcgt
[in the Rat DNA
Chips RG-U34B}
|
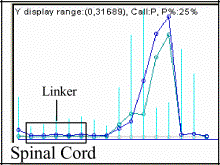
Disulfide Isomerase related prot. (Erp70)
|
Children's National Medical
Center Accesed Feb. 1, 2005.
|
Note:
The EcoRI-related palindromic linker ctcgtgccgaattcggcacgag
causes the drop of microarray expression to zero demonstrating its absence in
the tissues [dChip V.1.2 (9)]. Highlighted
in the second column in clear blue are the portions corresponding to the
palindromic linker, and in dark blue, the nucleotides exchanged to obtain the
second set or "mismatch" in Affymetrix'
probes (DNA-Chip).
The phenomenon of heterotranscription
Twelve
bases seem to be the minimum common denominator in order for the EcoRI palindromic linker to produce
artificial heterotranscripts such as the ones reported here and present in the Genbank.
The
most common palindromic flanks for the oligonucleotide ccgaattcgg are g and c, which give us
the longer oligonucleotide gccgaattcggc. Less frequent are the flanks c
and g to produce the second oligonucleotide cccgaattcggg,
with a similar effect on heterotranscription. This last palindromic sequence is
the one that we have in reference (1). The same palindromic sequence is present
in example 9 from Table 2 (Homo sapiens
X93499 for the RAB7 protein), in which we have fragments for more than two
genes attached together in the same strand, through the palindromic linkers ccccgaattcgggg
and gcccgaattcgggc
(12).
Table 2. Transcripts
with traces of an EcoRI related
linker and some related references.
# |
Accession/Organism |
Gene/Protein * |
Linker ** |
References *** |
|
1
|
U58090
Homo sapiens |
Cullin gene family member,
Hs-cul-4A |
aattcggcacgagctcgtgccgct
NSARARAA
|
Cell
85, 829-839. 1996.
|
|
2
|
U28831
Homo sapiens
|
Protein
immuno-reactive with anti-PTH polyclonal antibodies
|
gcacgagctcgtgccgat
ARARAD
|
Proc. Assoc. Am. Physicians 107, 296-305. 1995.
|
|
3
|
BC041619
Homo sapiens |
Protein
KIAA0404, for IMAGE:5923662 [R: hypoth. prot. MGC16044]
|
ccctcgtgccgaattcggcacgag
PSCRIRHE |
Proc. Natl. Acad. Sci. U.S.A. 99, 16899-16903. 2002. |
|
4
|
AF176705 Homo sapiens |
F-box protein FBX10 (PINX1) [R: vector] |
cctcgtgccgaattc PRAEF |
Curr. Biol. 9, 1180-2. 1999. |
5 |
X85792
Homo sapiens
|
Vpr binding
protein 1
|
tcgtgccgaattcggcacgag
SCRIRHE
|
Benichou et al. [Unpublished]; J Biol Chem. 277, 45091-8. 2002
|
6 |
AF151109
Homo sapiens
|
Putative
BRCA1-interacting protein (BRIP1)
|
ggcacgagctcgtgccgc
GTSSCR
|
Wang et al. BRCA1-interacting protein. [Unpublished]; Oncogene 19, 6152-8. 2000.
|
|
7
|
AF146697
Homo sapiens
|
FOXP1
|
aagaattcggcacgagct
KNSARA
|
Cancer Res. 61, 8820-8829. 2001.
|
|
8
|
NM_002342
Homo sapiens
|
Gene and 3' UTR for TNFR
superfamily, member 3 (LTBR)
|
gctcgtgccgaattc
|
Genomics 16, 214-218. 1993. [Curated by NCBI]
|
|
9
|
X93499
Homo sapiens
|
RAB7 protein,
GTP-binding [L: Dystroglycan 1. C: Rab7. R: Envelope glycoprotein]
|
ccccgaattcgggg
& gcccgaattcgggc
|
Biochem. Biophys. Res. Commun. 229, 887-890. 1996.
|
|
10
|
X82200
Homo sapiens
|
Gene and mRNA
for interferon-induced Staf50
|
gaattcggcacgagctc
|
J. Biol. Chem. 270, 14891-14898. 1995.
|
|
11
|
U31384
Homo sapiens
|
mRNA for G
protein gamma-11 subunit
|
ggcacgagctcgtgccg
|
J. Biol. Chem. 270, 21765-21771. 1995.
|
|
12
|
AF379619
Homo sapiens
|
Intron near
AB13, precursor mRNA
|
gaattcggcacgagct
|
van Roy and Staes. New
human gene family. [Unpublished].
|
|
13
|
AY245868
Homo sapiens
|
CDS for Aldehyde
oxidase-like protein (AOX2) pseudogene
|
aagaattcggcacgagca
LNSARA
|
Wright RM. Human aldehyde
oxidase. [Unpublished].
|
|
14
|
AF339764
Homo sapiens
|
mRNA from Fetal
liver spleen IMAGE:108721
|
gaattcggcacgagcggcacgagct
|
Genomics 79, 635-656. 2002.
|
|
15
|
U43527
Homo sapiens
|
5'UTR for Malignant
melanoma metastasis-suppressor (KiSS-1)
|
(ctct)15cctcgtgccgaattcggcacgag
|
J. Natl. Cancer Inst. 88, 1731-1737. 1996; Genomics 54, 145-148. 1998.
|
Notes: This
table presents the EcoRI related linker ctcgtgccgaattcggcacgag
as it appears in the Genbank for some human genes. To view the
rest of this Table 2 and the presence of the linker in other organisms, refer
to URL: http://www.oocities.org/plin9k/t2.htm
* Gene/Protein (Gene
symbol) [notes for sides of linker (L or R)]
** Linker and its Translation in Amino Acids as presented in the Genbank
*** Corresponding References According to the Genbank; closest related match
Abundance of sequences including the EcoRI-like palindromic linker
There
are thousands of sequences, including expressed
sequence tags (est) in the Genbank and in other nucleotide
databases that still contain artifacts, having as its common denominators, EcoRI palindromic linkers like the ones
described in this article.
The
palindromic linkers can be present in tandems, halves, or in different lengths;
being 12 to 24 bases its most common range. Artificial linkers have been found
even inside multiple coding regions, like the examples presented in the full
Table 2 (7). Examples of those linkers are frequently present outside the
coding region, i.e, in promoters,
like in:
1-) NR_001557 for H.
sapiens aldehyde oxidase 2 (AOH2) on chromosome 2, oligo gaattcggcacgagc
(13).
2-) NM_002342 H.
sapiens lymphotoxin beta-receptor (LTBR; member 3 of the TNFR superfamily),
oligo gctcgtgccgaattc (14).
Furthermore, those palindromic
linkers have been found also in the 5' region, i.e., in sequence U43527 for the human malignant melanoma
metastasis-suppressor KiSS-1, oligo (ctct)15cctcgtgccgaattcggcacgag
(15), and/or in the 3' region, i.e.,
sequence AY029161 for the Pin2-interacting protein
X1, oligo ctcgtgccgaattcggcac (16).
Of the
few submissions to the Genbank that
are explicitly reporting the presence of the EcoRI adapter, Hirama et al
(17) stands out, together with Inoue et
al (18) and Savas et al (19).
However, Inoue et al (18) considers
only the 8 first bases at the left flank as part of the sequence for the EcoRI-adapter. The effect of the
palindromic linker for Inoue's sequence may extend to at lest 16 bases
(sequence D83948 for S1-1 protein, oligo ggcacgagctcgtgccg) by an apparent
phenomenon of self-recombination and self-insertion inside the host-vector
interactions.
A
similar situation to what we see in Inoue's is presented by Savas et al (19), mentioning in their Genbank submission the first 6 bases
only as part of the EcoRI-adapter
(same in reference 1, but not in the submitted sequence L21934). In Savas'
reference (19) the linker-like effect may be extended about 20 bases (sequence
X78445 for Cytochrome P450 Cyp1-b-1, oligo gaattcggcacgaactcgtgc). Hirama et al (17) is the only one that
appropriately mentions a longer extension for the EcoRI-adapter, reporting it as being of 14 bases (sequence X56703
for the rearranged T-cell receptor alpha chain, oligo gaattcggcacgagct).
Chimaeras linked by the EcoRI-like palindrome seem to be resistant to enzymatic digestion
The
palindromic linkers persist in the sequences without being digested by the
enzymes. The discovery of mechanisms of resistance to the enzymatic digestion awaits
further study. However, it is evident that the most common palindromic linkers
match the identity of the EcoRI
adapter sequence gaattcggcacgag, which is used for the 5'UTR, and reported to
the Genbank, i.e., inside the sequence AI607511 as ctcgtgccgaattcggcacgag (similar
to acidic ribosomal phosphoprotein PO). In that sequence, it is indicated the
use of the vector pBluescript SK(-),
plus EcoRI, with the additional use
of the vector Uni-ZAP XR. These
methodologies, like the ones described in (1, 5, 6), may be promoting the
phenomenon of artificial linkage abundantly present in the Genbank.
The
rearrangement and splicing in a host-vector interaction resistant to EcoRI enzymatic digestion may be
explained by a phenomenon of self-hybridization performed by the linker (Figure
3), which could make it to appear as an appendage impossible to be grasped by
the digestive enzyme.
Figure 3.
Self-hybridization of the palindromic EcoRI-like
linker seems to block enzymatic digestion. Phenomenon also seen as 'a closing zipper', at both ends of longer
sequences, where two distant parts of the linker approach and stick together
producing a plasmid-like formation. [Examples 38-42, full Table 2 (7). Figure
obtained using the software from reference 20.]
CONCLUSIONS AND PERSPECTIVES
Stringent quality control on sequence databases is required
These
findings may contribute to a more stringent implementation of a quality control
within nucleotide databases, as well as the professional analysts of molecular
artifacts and related experiments, not to mention the rational human
engineering of pharmaceutical drugs and proteins. We must first tame, through
its controlled use, those palindromic linkers. Then, the use of sequence
quality control (to detect those molecular artifacts) can successfully be
applied. Improvements can be made in the design of Affymetrix microarrays, i.e.,
by removing the palindromic linker from human sequences such as AA557228,
AA113291, AI798671, AA864645, AA780435, W90032, AA810599, T52176, AA976510,
AI380906, AA535275, AI792166, T67559, L04270, U83598, D59474, T03148, T54342,
D59674, D59787, D80164, H90908, N80129, C14426, D59619, D80240, T56800, C14298,
W72424, IR1056496, T69555, D80337, C14227, C14407, C14344, D80210, etcetera.
Possible use of palindromes and its heterotranscripts to reverse
hereditary diseases
Another
possibility is the use of palindromes for the attachment of two different genes
(genetic modules) for the engineering of therapeutic proteins, i.e., for the engineering of antiobesity
treatments that may be customized and prescribed according to the particular
metabolism of each person (21).
Artificial vectors for gene therapy may produce heterotranscription
It is
possible that palindromic linkers like the ones reported in this article, or
using other linkers, may be produced by artificial vectors for gene therapy
also. If this is the case, an unsuspected side effect for humans may result. To
date, those linkers are still present in those thousands of artificial
sequences submitted to the Genbank,
and are an allegory of things that can be prevented in human gene therapy. This
article has been written to raise the awareness to the scientific community on
the presence of these thousands of artificial hetero-transcripts by current
methodologies producing the sequences reported to the Genbank as well as the vectors used in gene therapy.
Published and submitted heterotranscripts are still awaiting its correction
In
doing an analysis of the sequences presented as chimerical mRNAs in Table 4 of
reference (22), I also found in that article the EcoR1-related palindromic linker present inside the sequence
AY029161 (16), already seen. In that sequence, there is a fragment for a
putative human tumor suppressor LPTL, AF418553, originated in the human
chromosome 8, and linked with a fragment for phosphohistidine phosphatase
PHP14, NM_014172 originated in the human chromosome 9.
Until
the submission of this article, none of the sequences presented here had been
corrected in the Genbank. Could this
be because they have not been determined as artifact products to date?
Another
sequence in reference (22) that contained the EcoR1 related palindromic linker was BC000519, identified as an
artifact and has already been removed from the Genbank.
A possible transposon-like removal of
palindromic linkers seem to have happened on other chimeras
Awaiting
correction are thousands of sequences like the ones presented in Table 2 (7).
Sequences like AF176705 for the human F-box protein FBX10 (23), and Z28355 from
the atrium heart (24), are both containing fragments of the vector and of the
linker. Another sequence, HTCBYB08, contains fragments of the vector only,
lacking the linker. Experiments were also done with such sequences lacking any
evidence of a palindromic linker, also without obtaining amplified products by
RT-PCRs (unpublished results). Both
of the sequences Z28355 and HTCBYB08 exhibit traces of the same cloning vector,
vector from the sequence X52324 (ARBLSKM), the pBluescript SK(-) vector (25-27). A transposon-like self-removal of
the palindromic linker may be one way in which many heterotranscripts appear to
be lacking the linker, as in the sequence HTCBYB08. This hypothesis also needs
to be experimentally evaluated. If this is the case, palindromic linkers may be
used for a possible artificial reversal of mutations in vivo (28).
On the
other hand, heterotranscripts lacking the linker, and present in Affymetrix' microarrays, may show two
different expression profiles, for example, a zero expression on one side while
a contrasting higher expression on the other side (data not shown). A similar pattern is present when non-expressed
introns are included within Affymetrix'
probes side by side with sequences expressed by the exons, as in Figure 1 from
reference (2).
Final comment
The key
palindrome described in this article has already been introduced in reference
(2) as CTCGTGCCGAATTCGGCACGAG and it
has been reported elsewhere (2, 3, 29), also.
ACKNOWLEDGMENTS
Tracy
Lynn Duncan helped in preparing this review and supported me during the last
two years.
REFERENCES
1. Li
BL, Li XL, Duan ZJ, Lee O, Lin S, Ma ZM, Chang CC, Yang XY, Park JP, Mohandas
TK, Noll W, Chan L & Chang TY (1999) Human Acyl-CoA:Cholesterol
Acyltransferase-1 (ACAT-1) gene organization and evidence that the 4.3-kilobase
ACAT-1 mRNA is produced from two different chromosomes. J. Biol. Chem. 274: 11060-11071.
2. Castro-Chavez
F (2004) Microarrays, antiobesity and the liver. Ann. Hepatol. 3: 137-145. [http://www.medigraphic.com/pdfs/hepato/ah-2004/ah044c.pdf]. Accessed Oct. 1, 2005.
3.
Castro-Chavez F (2004) Some Implications for the Study of Intelligent Design
Derived from Molecular and Microarray Analysis. PCID 3.1.7 (serial online), November, Issue 3. [http://www.iscid.org/pcid/2004/3/1/pcid_contents_2004_3_1.php].
Accessed Oct. 1, 2005.
4.
International Human Genome Sequencing Consortium (2004) Finishing the
euchromatic sequence of the human genome. Nature
431: 931-45.
5. Yang
L, Lee O, Chen J, Chen J, Chang CC, Zhou P, Wang ZZ, Ma HH, Sha HF, Feng JX,
Wang Y, Yang XY, Wang L, Dong R, Ornvold K, Li BL & Chang TY (2004) Human
acyl-coenzyme A:cholesterol acyltransferase 1 (acat1) sequences located in two
different chromosomes (7 and 1) are required to produce a novel ACAT1 isoenzyme
with additional sequence at the N terminus. J.
Biol. Chem. 279: 46253-62.
6. Chang
CC, Huh HY, Cadigan KM & Chang TY (1993) Molecular cloning and functional
expression of human acyl-coenzyme A:cholesterol acyltransferase cDNA in mutant
Chinese hamster ovary cells. J. Biol.
Chem. 268: 20747-55.
7. Castro-Chavez
F (2005) Full version of Table 2. Website. [http://www.oocities.org/plin9k/t2.htm].
Accessed Oct. 1, 2005.
8. Castro-Chavez
F, Yechoor VK, Saha P, Martinez-Botas J, Wooten E, Sharma S, O'Connell P,
Taegtmeyer H & Chan L (2003) Coordinated upregulation of oxidative pathways
and downregulation of lipid biosynthesis underlie obesity resistance in
Perilipin knockout mice. A microarray gene expression profile. Diabetes 52: 2666-2674. [http://diabetes.diabetesjournals.org/cgi/content/full/52/11/2666].
Accessed Oct. 1, 2005.
9. Li C & Wong WH (2001) Model-based analysis of oligonucleotide arrays: expression index computation and outlier
detection. Proc. Natl. Acad. Sci. U.S.A. 98: 31-36. [http://biosun1.harvard.edu/complab/dchip]. Accessed Oct. 1, 2005.
10. Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W & Lipman DJ (1997) Gapped BLAST and
PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 25: 3389-3402
[http://www.ncbi.nlm.nih.gov/BLAST]. Accessed Oct. 1, 2005.
11. Castro-Chavez F (2004) El Incremento Coordinado de las Rutas Metabolicas Oxidativas y la Disminucion en la
Biosintesis de Lipidos Caracterizan la Resistencia a la Obesidad en el Raton Artificialmente Carente de Perilipina.
Doctoral thesis, Universidad de Guadalajara, Guadalajara, Jalisco, Mexico.
[http://www.oocities.org/plin9k/tesis-fdocc.zip]. Accessed Oct. 1, 2005.
12. Vitelli R, Chiariello M, Lattero D, Bruni CB & Bucci C (1996) Molecular cloning and expression analysis of the human
Rab7 GTP-ase complementary deoxyribonucleic acid. Biochem. Biophys. Res. Commun. 229: 887-890.
13.
Terao M, Kurosaki M, Marini M, Vanoni MA, Saltini G, Bonetto V, Bastone A,
Federico C, Saccone S, Fanelli R, Salmona M & Garattini E (2001)
Purification of the aldehyde oxidase homolog 1 (AOH1) protein and cloning of
the AOH1 and aldehyde oxidase homolog 2 (AOH2) genes. Identification of a novel
molybdo-flavoprotein gene cluster on mouse chromosome 1. J. Biol. Chem. 276: 46347-63.
14. Matsuda A, Suzuki Y, Honda G, Muramatsu S, Matsuzaki O, Nagano Y, Doi T, Shimotohno K, Harada T, Nishida E,
Hayashi H & Sugano S (2003) Large-scale identification and characterization of human genes that activate NF-kappaB and
MAPK signaling pathways. Oncogene 22: 3307-3318.
15. West
A, Vojta PJ, Welch DR. & Weissman BE (1998) Chromosome localization and genomic
structure of the KiSS-1 metastasis suppressor gene (KISS1). Genomics 54: 145- 148.
16. Zhou XZ & Lu
KP (2001) The Pin2/TRF1-interacting protein PinX1 is a potent telomerase
inhibitor. Cell 107: 347-359.
17.
Hirama T, Takeshita S, Matsubayashi Y, Iwashiro M, Masuda T, Kuribayashi K,
Yoshida Y & Yamagishi H (1991) Conserved V(D)J junctional sequence of
cross-reactive cytotoxic T cell receptor idiotype and the effect of a single
amino acid substitution. Eur. J. Immunol.
21: 483-488.
18.
Inoue A, Takahashi KP, Kimura M, Watanabe T & Morisawa S (1996) Molecular
cloning of a RNA binding protein, S1-1. Nucleic
Acids Res. 24: 2990-7.
19.
Savas U, Bhattacharyya KK, Christou M, Alexander DL & Jefcoate CR (1994)
Mouse cytochrome P-450EF, representative of a new 1B subfamily of cytochrome
P-450s. Cloning, sequence determination, and tissue expression. J Biol Chem. 269: 14905-11.
20. Zuker
M. (2003) Mfold web server for
nucleic acid folding and hybridization prediction. Nucleic Acids Res. 31: 3406-15.
21. Qi L, Shen H,
Larson I, Schaefer EJ, Greenberg AS, Tregouet DA, Corella D & Ordovas JM.
(2004) Gender-specific association of a perilipin gene haplotype with obesity
risk in a white population. Obes. Res.
12: 1758-65.
22.
Romani A, Guerra E, Trerotola M & Alberti S (2003) Detection and analysis
of spliced chimeric mRNAs in sequence databanks. Nucleic Acids Res. 31, e17: 1-8.
23.
Winston JT, Koepp DM, Zhu C, Elledge SJ & Harper JW (1999) A family of
mammalian F-box proteins. Curr. Biol.
9: 1180-1182.
24. Auffray
C, Behar G, Bois F, Bouchier C, Da Silva C, Devignes MD, Duprat S, Houlgatte R,
Jumeau MN, Lamy B (1995) IMAGE:
molecular integration of the analysis of the human genome and its expression. C. R. Acad. Sci. III, Sci. Vie. 318:
263-72.
25.
Short JM, Fernandez JM, Sorge JA & Huse WD (1988) Lambda ZAP: a bacteriophage lambda expression vector with in vivo excision properties. Nucleic Acids Res. 16: 7583-7600.
26.
Alting-Mees MA & Short JM (1989) pBluescript
II: gene mapping vectors. Nucleic
Acids Res. 17: 9494.
27. Alting-Mees MA, Sorge JA & Short
JM (1992) pBluescriptII:
multifunctional cloning and mapping vectors. Meth. Enzymol. 216: 483-495.
28.
Hirschhorn R (2003) In vivo reversion
to normal of inherited mutations in humans. J.
Med. Genet. 40: 721-728. [http://jmg.bmjjournals.com/cgi/content/full/40/10/721]
Accessed Oct. 1, 2005.
29. Castro-Chavez F (2005) Genes involucrados en la
susceptibilidad a la obesidad y los genes anti-obesidad. In: Mendez-Sanchez
N (Editor), OBESIDAD. Epidemiologia,
Fisiopatologia y Manifestaciones Clinicas. 2nd Ed. Manual
Moderno. Mexico (in press).
////////////////////////////////////////////////////////////////
Notes:
The PDF of this work 'Palindromati' can be found at:
http://www.iscid.org/boards/ubb-get_topic-f-6-t-000582.html
Mendelian Bioengineering:
http://www.oocities.org/plin9k/bioeng.htm
http://www.oocities.org/plin9k/limiting-species.htm
Links to the Biggest Forensics Thus Far.
Blog 'Research on Intelligent Design':
The Index # 1 for my Blog 'Research on Intelligent Design':
The Index # 2 for my Blog 'Research on Intelligent Design':
The Index # 3 for my Blog 'Research on Intelligent Design':
Tasters of the Word (YouTube), videos recientes: "Astronomía y Nacimiento de Jesucristo: Once de Septiembre Año Tres A.C.", "Estudio sobre Sanidades" (en 20 episodios), "Jesus Christ, Son or God?":http://www.youtube.com/1fertra
Tasters of the Word (the blog, with: "Astronomy and the Birth of Jesus Christ"):http://fertra1.blogspot.com