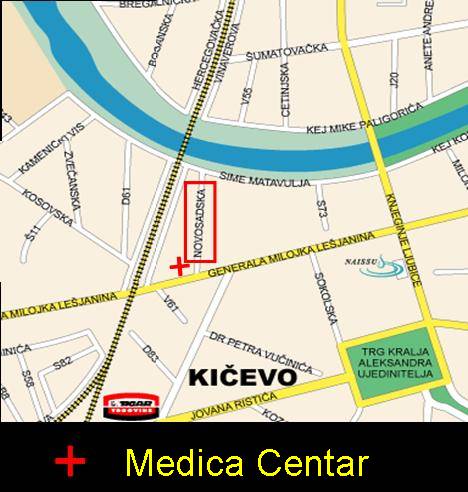
Pocetna Engleski Kontakt Kako da nas pronadjete
{kind=link}
Dobrodosli na sajt medica centra
Designed on October, 2008 by Ivan Bubanovic
 |
Download |
 |
|
 |
Tema meseca:
|
CITOKINI I FUNKCIJA
TESTISA
ULOGA CITOKINA U
FUNKCIJI TESTISA
Dokazi o povezanosti
testikularne funkcije i inflamatornih oboljenja mogu se naci u radovima starijeg
i novijeg datuma. Poznato je da su inflamatorna oboljenja udruzena sa znacajnim
skokom tkivne i serumske koncentracije proinflamatornih citokina koji mogu
znacajno uticati na proces steroidogeneze i spermatogeneze. Saznanja o ulozi
citokina u procesima steroidogeneze i spermatogeneze za vreme patoloskih stanja
nedvosmisleno su ukazala na ulogu ovih faktora u fizologiji testikularne
funkcije. Citokini se u tom smislu pojavljuju kao faktori rasta, diferencijacije
i apoptoze. Osim toga, citokini mogu znacajno uticati na funkcionisanje osovine
hipotalamus-hipofiza-testisi
(H-H-T) i na taj nacin dopuniti svoje efekte na reproduktivnu funkciju testisa.
Spermatogeneza je
kontinuirani proces koji se odvija iza krvnotestikularne barijere.
Sekvestriranost ovog procesa je najverovatnije nametnula razvijanje kontrolnih
mehanizama "zastite" spertogenog loze (epitela) od sopstvenog
imunskog sistema koji pocivaju na parakrinim i autokrinim faktorima. Pored neosporne
uloge FSH i androgena kao regulatornih faktora, kontrolni
mehanizmi spermatogeneze
najvecim delom pocivaju na ulozi parakrinih faktora kao sto su faktori rasta i
citokini koji uglavnom poticu iz epitela reproduktivnog kanala, makrofaga,
imunskih celija, Leydigovih i Sertolijevih celija. Ukoliko
zelite da saznate vise o povezanosti muskog steriliteta sa autoimnskim
mehanizmima - kliknite ovde.
IMUNO-ENDOKRINA
KONTROLA FUNKCIJE LEYDIGOVIH CELIJA
Leydigove celije su
centralno mesto produkcije androgena. Uzimajuci u obzir cinjenicu da Leydigove
celije nisu izolovane krvno-testikularnom barijerom,
nije tesko zakljuciti da faktori kontrole androgeneze Leydigovih celija mogu
poticati iz samog intersticijuma testisa kao i ostalih endokrinih zlezdi.
Biosinteza testosterona kao najvaznijeg androgena Leydigovih celija primarno je
pod kontrolom LH i usko je povezana sa funkcionisanjem citave H-H-T osovine.
Posle vezivanja sa specificnim receptorom LH stimulise sintezu cAMP u
Leydigovim celiama. Nakon ovog intracelularnog signala sledi aktivacija cAMP
zavisnih protein kinaza. cAMP stimulise androgenezeu putem dva mehanzma: (i)
mobilizacija i aktivacija transporta holesterola, sto se dogadja u intervalu od
svega nekoliko minuti i (ii) dugotrajna ekspresija gena steroidogenetskih
enzima. Ovaj efekat se ocitava tek nekoliko sati nakon pristizanja LH signala
(1).
Danas je vecina autora
saglasna u misljenju da proinflamatorni citokini snazno inhibisu gonadotropinima i cAMP-om
posredovanu androgenezu u Leydigovim celijama. Citokini koji su najcesce
udruzeni sa bakterijskim i virusnim zapaljenjskim oboljenjima su IL-6, TNF,
IL-1, IL-2 i IFN-g.
Svi navedeni citokini, izuzev IL-6, inhibisu cAMP-om posredovanu androgenezu u
Leydigovim celijama, kao i ekspresiju enzima neophodnih za proces androgeneze
(2,3).
Oplodjenje nije "sudar"
permatozoida i jajna celije vec mnogo vise od toga.
INTERAKCIJA
MAKROFAGA I LEYDIGOVIH CELIJA
Kao sto je vec receno, u imunoloskom smislu
testis predstavlja "priviligovani organ". Iz ovih razloga alotransplantat ili
ksenotransplantat prezivljavaju duze ako se implantiraju u tkivo testisa u
odnosu na transplantate koji su implantirani u npr. poprecnoprugastu muskulaturu
(u imunoloskom smislu "nepriviligovan organ" sto znaci da nije osepariran tzv.
krv-organ barijerom) (4).
Testikularni makrofazi
cine oko 25% celija intersticijuma testisa. Makrofazi intersticijuma testisa
(MIT) su veoma slicni makrofazima ostalih tkiva, eksprimiraju MHC
molekule klase
II, mogu da vrse fagocitozu i eksprimiraju Fc receptor. Nakon antigenske
stimulacije MIT sekretuju IL-1 i ostale proinflamatorne citokine, osim toga
stimulisani MIT povecavaju broj eksprimiranih kostimulatornih molekula (5,6).
Urodjeni nedostatak MIT kod nekih sojeva miseva dovodi do teskih poremecaja u
razvoju testisa, funkcionisanju Leydigovih celija i spermatogeneze. Leydigove
celije i makrofazi intersticijuma testisa pokazuju blisku anatomsku povezanost,
koja nedvosmisleno ukazuje na aktivnu komunikaciju izmedju ovih celija. U osnovi
ove komunikacije najverovatnije su sadrzani neki od regulatornih mehanizama
Leydigovih celija (6). Na Leydigovim celijama testisa pacova
otkrivene su tzv. "membransko-citoplazmatske
digitacije" koje nalezu na "membarnsko-citoplazmatske" invaginacije
makrofagalnih celija intersticijuma testisa. Ovaj vid "fizicke"
povezanosti MIT i Leydigovih celija ukazuje i na njihovu
blisku funkcionalnu povezanost (6). Osim toga, paralelno sa
Leydigovim celijama, MIT podlezu morfoloskim i funkcionalnim promenama za vreme
razvoja testisa i to narocito u vreme puberteta (7). Leydigove celije
eksprimiraju receptore za mnoge citokine i to pretezno one citokine koje luce
MIT i ostale imunske i neimunske celije intersticijuma testisa (8).
Nedavno su otkrivene
neke specificnosti MIT u odosu na makrofage ostalih tkiva i pretpostavlja se da
ove specificnosti cine osnovu aktivne komunikacije sa Leydigovim celijama. Iako
je dokazano da MIT mogu sekretovati citokine kao sto su IL-1, IL-6, TNF-a,
GM-CSF i sl., sposobnost njihovog odgovora na antigensku stimulaciju je znacajno
niza od npr. odgovora peritonealnih makrofaga. Osim toga, MIT odgovaraju na
antigensku stimulaciju nesto drugacijim “setom” citokina u odnosu na
peritonealne makrofage (9). Neuobicajena pojava za makrofage ostalih tkiva, je
da MIT eksprimiraju receptore za FSH, kao i da FSH ima uticaja na citokinsku
aktivnost MIT (10). Ekspresija kostimulatornih liganda B7-1 i B7-2 je znatno
slabija kod MIT nego kod makrofaga ostalih tkiva, sto ukazuje na cinjenicu da
MIT mogu indukovati T celijsku anergiju ili cak deleceiju specificnih klonova T
limfocita (11). Ove specificnosti MIT pomazu u odrzavanju
imunske tolerancije u odnosu na testikularni epitel, ali u isto vreme to znaci
da poremecaj funkcije MIT moze biti odgovoran za autoimunski orhit, ostecenje
spermaticne loze, slabije parametre spermograma i konacno sterilitet muskaraca.
Ukoliko zelite da saznate vise o povezanosti muskog steriliteta sa autoimnskim
mehanizmima - kliknite ovde.
Citokini kao sto su
IL-1 i TNF-a,
sekretovani od strane MIT, mogu dostici veoma visoke koncentracije u
mikrookolini Laydigovih celija i na taj nacin znacajno doprineti usporavanju
androgeneze. Osim MIT, IL-1 i IL-6 izlucuju i Sertolijeve celije koje takodje
mogu doprineti mehanizmu citokinske kontrole androgeneze (12). Istrazivanja na
animalnim modelima su pokazala da injekcija lipopolisaharida gram negativnih
bakterija (LPS) direktno aplikovana u testis aktivira MIT, dovodi povecanja
produkcije proinflamatornih citokina i znacajnog pada produkcije testosterona,
kao i
njegove serumske koncentracije (6,12). Osim parakrine regulatorne funkcije
citokina u kontroli androgeneze, dokazana je sekrecija IL-1, TNF-a
i IL-6 iz samih Leydigovih celija, sto ukazuje na postojanje autokrinih
mehanizama citokinske kontrole androgeneze o kojoj se vrlo malo zna (12,13).
EFEKTI CITOKINA NA
FUNKCIJU LEYDIGOVIH CELIJA
Iako jos uvek postoje
neka neslaganja oko efekta IL-1 na androgenetski aparat Leydigovih celija,
vecina autora se slaze da IL-1 snazno inhibise LH/hCG i/ili cAMP stimulisanu
sintezu i sekreciju testosterona (13). IL-1 posredovana inihibicija androgeneze
je najvecim delom uslovljena blokiranjem aktivacije inducibilnih citohrom P450
enzima cija je uloga u sintezi polnih hormona presudna, dok ekspresija bazalnih
vrednosti konstitutivnih izoformi ovih enzima nije zavisna od IL-1 (14).
TNF-a
je jedan od najpotentnijih citokina kada je u pitanju njihov uticaj na
intenzitet androgeneze. Iako neki autori smatraju da TNF-a
stimulise steroidogenezu, vecina autora ubraja TNF-a
u snazne inhibitore produkcije testosterona. Istrazivanja sprovedena na
animalnim modelima, kao i in vitro istrazivanja na izolovanim Leydigovim
celijama i MA-10 tumorskim Leydigovim celijama ukazuju na to da TNF-a
inhibise androgenezu cak i onda kada je ona stimulisana putem hCG. Osnovni
mehanizam inhibitorne akcije TNF-a
je supresija ekspresije gena steroidogenetskih enzima i snizavanje cAMP u
Leydigovim celijama. TNF-a
kao i IL-1 nema uticaja na bazalnu ekspresiju steroidogenetskih enzima (15).
IL-2 snazno stimulise
osovinu H-H-nadbubrezne zlezde. Rezultati ove stimulacije se ogledaju u
povecanju sekrecija ACTH i glukokortikosteroida nadbubreznih zlezdi. Povecane
vrednosti ovih hormona znacajno suprimiraju izlucivanje gonadotropina, kao i
androgenezu u Leydigovim celijama. IL-2 pokazuje i direktne inhibitorne efekte
na androgenezu u Leydigovim celijama. Slicno IL-1 i TNF-a,
IL-2 inhibise androgenezu na nivou enzimskog kompleksa P450 (16). Pacijenti
oboleli od raka koji su leceni visokim dozama IL-2 pokazuju veoma niske serumske
koncentracije testosterona, bez obzira na starosnu dob i lokalizaciju primarnog
procesa (17). Uzimajuci u obzir da povecana IL-2 aktivnost rezultuje skokom serumskih i
tkivnih koncentracija ostalih proinflamatornih citokina kao sto su IL-1, TNF-a
i IFN-g,
jedan deo inhibitornih efekata IL-2 na androgenezu svakako se ostvaruje
posredovanjem ostalih proinflamatornih citokina (16,17,18).
Interferoni (IFN-a,
IFN-b
i IFN-g)
nemaju direktnog uticaja na hipofizu i izlucivanje gonadotropina, ali efikasno
suprimiraju sintezu testosterona u Leydigovim celijama cak i onda kada je ona
stimulisana prisustvom hCG. Mehanizam akcije IFN se zasniva na blokiranju
transporta holesterola u mitohondrije Leydigovih celija, kao i na supresiji
aktivnosti pojedinih enzima iz kompleksa P450 (19). Istrazivanja sprovedena na
ljudima su pokazala da su virusne infekcije najcesce udruzene sa porastom
serumske i tkivne koncentracije IFN i znacajnim padom serumske koncentraceije
testosterona (20). Osim klasicnih proinflamatornih citokina, inhibitorne ucinke
na androgenetsku aktivnost Leydigovih celija pokazuju i neki antiinflamatorni
citokini poput TGF (a, b1, b2
i b3).
TGF‑b
inhibise androgenezu na nivou LH receptora, smanjujuci njihov broj i osetljivost
za LH (21).
EFEKTI ANDROGENA NA
IMUNSKI SISTEM
Nekoliko znacajnih
studija sprovedenih na animalnim i humanim modelima potvrdile su zapazanje da
jedinke muskog pola imaju znacajno slabiji imunski odgovor od jedinki zenskog
pola. Najverovatnije iz ovih razloga incidenca oboljevanja od infektivnih
bolesti i malignih tumora je znatno veca medju pripadnicima muskog pola. Nasuprot
ovome, incidenca autoimunskih oboljenja je veca kod pripadnica zenskog pola.
Najveci broj autora se slaze oko pretpostavke da su razlike u miljeu steroidnih
polnih hormona i njihovom ritmu emitovanja odgovorne za znacajne razlike u
kvalitetu i kvantitetu imunskog odgovora izmedju jedinki muskog i zenskog pola
(22,23).
Veza izmedju serumskog
nivoa testosterona i tezine reumatoidnog artritisa, jasno je ukazala na znacajne
imunsupresivne efekte ovog hormona. Naime, osobe sa niskim nivoom sermskog
testosterona znatno cesce oboljevaju od reumatoidnog artritisa i to u tezim
formama. In vitro istrazivanja su pokazala da testosteron snazno inhibise
produkciju IL-1 od strane monocitnih celija periferne krvi (23,24).
Dihidrotestosteron inhibise produkciju IL-4, IL-5 i IFN-g
cak i onda kada su periferni limfociti stimulisani mitogenima sto ukazuje na
regulatornu ulogu androgena u procesu razvijanja imunske reakcije (25).
Androgeni su poznati i kao faktori involucije timusa uslovljavajuci gubitak
mase timusa na racun smanjenja broja limfocita u kortikalnoj zoni. Nasuprot
ovome, hiperplazija timusa i uvecanje njegove mase je uobicajeni nalaz kod
kastriranih zivotinja i ljudi (26).
ULOGA CITOKINA
U SPERMATOGENEZI
Kontrola procesa
spermatogeneze pored gonadotropina i androgena ukljucuje i parakrine faktore
kao sto su faktori rasta i citokini. Proces spermatogeneze sisara se sastoji
iz tri faze: mitoticka proliferacija spermatogonija, mejoticka redukcija
broja hromozoma i maturaciona faza do zrelih spermatozoida. U svakoj od
navedenih faza citokini imaju ulogu faktora rasta i/ili faktora
diferencijacije i maturacije spermatozoida (1,22).
Sekrecija IL-1a
od strane Sertolijevih celija je u tesnoj vezi sa njihovom fagocitnom
funkcijom. Naime, produkcija i sekrecija IL-1a
od strane Sertolijevih celija je uvek povecana kada postoji fagocitna
aktivnost ovih celija (5,6,8). Seminiferni epitel kao i Sertolijeve celije
sekretuju IL-6 kao odgovor na prisustvo FSH, mada je dokazana i bazalna
produkcija ovog citokina od strane navedenih celija (27). Egzogeni IL-6
inhibise spermatogenezu i to uglavnom mehanizmom inhibicije sinteze DNK u
mejotickoj fazi ciklusa (28).
De i sar. (29) su
pokazali da spermatociti iz faze pahiten i okrugle spermatide
eksprimiraju mRNA za TNF-a,
dok Sertolijeve celije eksprimiraju receptore za TNF-a
(8,29). Ovi podaci bi mogli ukazati na aktivnu citokinsku komunikaciju
izmedju celija spermatozoidne loze i Sertolijevih celija. Celije ranih faza
spermatogeneze ne samo da sintetisu TNF-a
vec i eksprimiraju receptor za TNF-a.
Aktivacija ovih receptora po dozno zavisnom modelu suprimira ekspresiju Fas
liganda i na taj nacin odlaze apoptozu germinativnih celija sprematozoidne
loze (30).
Produkcija IFN-a
i b
od strane Sertolijevih celija je takodje potvrdjena, dok IFN-g
produkuju samo spermatide u ranoj fazi razvoja (31). Medjutim, tretman miseva
IFN-g
dovodi do atrofije testisa, smanjenja zapremine ejakulata, abnormalne
morfologije spermatozoida, smanjenja broja spermatozoida u ejakulatu i
smanjenom fertilnoscu tretiranih miseva (32).
Iz ovih razloga vrlo je razumno aplikovati antiinflamatornu terapiju u cilju
poboljsanja parametara spermograma. Postoje studije koje potvrdjuju
opravdanost ovakvog pristupa u lecenju slabih parametara spermograma.
Ukoliko zelite da saznate vise o tome -
kliknite
ovde.
Koncentracija EGF
u serumu infertilnih muskaraca je znacajno niza od koncentracije ovog
citokina u serumu fertilnih muskaraca. Serumska koncentracija EGF je u
direktnoj korelaciji sa parametrima spermograma kao sto su zapremina
ejakulata, pokretiljivost i broj spermatozoida. Osim toga, utvrdjena je i
korelacija koncentracije serumskog EGF sa serumskom koncentracaijom hormona
kao sto su testosteron, LH i FSH (vidi grafikon) (33). Istrazivanja sprovedena na
tzv. knock out EGF deficijentnim misevima su pokazala kljucnu ulogu
EGF za proces spermatogeneze. Naime, EGF deficijentni misevi su infertilni,
zbog potpunog izostanka spermatogeneze (34). Isti autori su na potpuno
drugacijem eksperimentalnom modelu ponovo dokazali neophodnost prisustva EGF
za odvijanje spermatogeneze. U eksperimentu sa pacivima kojima su hirurskim
putem odstranjene pljuvacne zlezde dokazana je znacajno niza koncentracija
serumskog EGF uz ozbiljno kompromitovanje spermatogeneze. Tretman ovih
zivotinja EGF popravlja spermatogenezu skoro do fizioloskih granica (34).
Ovi interesantni podaci su otvorili prostor za nove spekulacije oko mogucih
imunopatoloskih mehanizama orhitisa i muskog steriliteta
kao komplikacije parotitisa (zauske).
Spermatogeneza je
u istoj meri zavisna od prisustva Platelet-derived growth factor
(PDGF) kao i od EGF. Kod PDGF deficijentnih miseva je nadjeno znacajno
smanjenje volumena testisa, drasticno smanjenje broja Leydigovih celija i
zastoj u procesu spermatogeneze. Rekonstrukcija soja PDGF deficiejntnih
miseva ukrstanjem sa divljim misevima rezultuje normalizacijom produkcije
PDGF, normalnim volumenom testisa i broja Leydigovih celija, kao i normalnim
odvijanjem spermatogeneze (35).
AUTOIMUNSKA
OBOLJENJA TESTISA
Testisi svoj
razvoj dovrsavaju za vreme postnatalnog zivota, a spermatogeneza pocinje za
vreme puberteta, tako da iz ovih razloga imunski sistem nije u prilici da
"upozna" novonastale celije, njihove antigene i faktore regulacije ovih
procesa, kao self antigene. Bez obzira na cinjenicu da fetalna
imunska tolerancija (IT) nije uspostavljena na mnoge antigene adultnog
testisa, tkiva testisa proces spermatogeneze nije ugrozen od strane
imunskog sistema zbog postojanja barijere krv-testis, koja prakticno
izoluje ova tkiva od nadzora imunokompetentnih celija. Celularnu osnovu
barijere krv-testis cine Sertolijeve i peritubularne celije koje
eksprimiraju veliki broj adhezionih molekula koji omogucavaju veoma jaku i
kompaktnu vezu izmedju ovih celija. Traumatski, zapaljenjski i infektivni
procesi na testisima mogu da naruse integritet barijere krv-testis i
otpocnu ciklus autoimunskog oboljenja testisa, koje neminovno vodi u
ostecenje procesa spermatogeneze i sterilitet (4,21). O opasnostima
imunizacije spermatozoidnim antigenima najbolje govori podatak da
imunizacija alogenim, pa cak i ksenogenim spermatozoidima kod
eksperimentalnih zivotinja moze dovesti do razvijanja antispermatozoidnih
antitela i auotimunskog orhitisa. Teuscher i sar. (36) opisuju uspesnu
imunizaciju zdrebadi spermatozoidima i adjuvansom, koja je proizvela dobar
antispermatozoidni odgovor sa znacajnim titrom antispermatozoidnih antitela
uglavnom IgG klase (36). Kod ljudi je primeceno da veliki procenat
vazektomisanih pacijenata pokazuje relativno visok titar antispermatozoidnih
antitela. Pretpostvlja se da do senzibilizacije na vlastite spermatozoide
dolazi zbog ekstravazacije spermatozoida tokom vazektomije (37).
Ukoliko zelite da saznate vise o tome - kliknite
ovde.
Danas se smatra da
kljucnu ulogu u formiranju krv-testis barijere imaju tzv. jake
inter-Sertolijeve veze (inter-Sertoli
tight junctions
- TJs). Ekspresija TJs proteina je najvecim delom pod kontrolom citokina kao
sto su TGF-b1,2
i 3.
U tom smislu TGF-b3
se najcesce pominje kao snazan supresor ekspresije TJs proteina i kao takav
igra vaznu ulogu imunopatoloskim mehanizmima bolesti testisa (38).
Osim barijere
krv-testis, dokazan je citav niz drugih mehanizama koji doprinose boljoj
zastiti procesa spermatogeneze i celija koje ucestvuju u spermatogenezi od
potencijalnog razvijanja autoimunske agresije. Sainio-Pollanen i sar. (11)
su dokazali potpuno odsustvo B7-1 (CD80) i B7-2 (CD86) pozitivnih
imunokompetentnih celija u testisima normalnih BALB/c miseva, dok je u
testisima NOD miseva dokazan znacajan broj CD80 i CD86 pozitivnih celija.
Najverovatnije iz ovih razloga NOD misevi spontano razvijaju autoimunski
orhitis i druge autoimunske bolesti, dok kod BALB/c miseva to nije slucaj.
Ovi autori sugerisu zakljucak da izostanak ekspresije kostimulirajucih
liganda B7-1 i B7-2 na MHC eksprimirajucim celijama dovodi do anergije T
helper CD4+ (Th) limfocita i do izostanka aktivacije
auto-citotoksicnih CTL. Nausoprot ovome, B7 pozitivne antigen prezentujuce
celije (APCs), aktivirane Th celije i CTL su dominantne celijske populacije
zapaljenjskog infiltrata u testisima NOD miseva (11). Ovi podaci ukazuju na
cinjenicu da u tkivima testisa postoje populacije makrofaga i dendriticnih
celija koje uspesno presecaju puteve aktivacije Th celija i dalju kaskadu
razvijanja celularnog i humoralnog autoimunskog odgovora. U isto vreme, ovaj
mehanizam uspesno objasnjava rezultate produzenog prezivljavanja alografta
implantiranog u tkivo testisa ili epididimisa (39).
U epitelu
epididimisa se mogu dokazati tzv. intraepitelni leukociti (IEL), cija se
koncentracija ne razlikuje od koncentracije IEL u drugim epitelinim tkivima.
Vecina studija svedoci o tome da CD8+ IEL mogu efikasno
suprimirati aktivnost CD4+ IEL, cime doprinose razvijanju
fenomena "hladjenja sluzokoza", sto je veoma vazno za odrzavanje
propustljivosti sluzokoza i njihove normalne funkcije. Osim toga, u epitelu
epididimisa dokazana je znacajna aktivnost TGF-b,
IL-4 i IL-10 sekretujucih limfocita. Smatra se da su opste zakonitosti
funkcionisanja MALT primenljive i na sluzokozu epididimisa, ukljucujuci i
mehanizme bystander tolerancije (40).
Komponente
komplementa C3 i C9 nisu dokazane u semenoj tecnosti ljudi, dok je prisustvo
C3 komponente komplementa u semenoj tecnosti zeca dokazano, ali njegova
koncentracija iznosi svega 25% od serumske koncentracije ove komponente
komplementa. U isto vreme inhibitori komplementa kao sto su CD46, CD55 i
CD59 dokazani su na spermatozoidima u seminoj plazmi i sekretu epididimisa
(41).
Smatra se da je za
inicijaciju autoimunskog oboljenja testisa neophodna aktivacija makrofaga,
povecanje koncentracije cirkulisuceg IFN-g
i aktivaciju CD4+ limfocita, koji su odgovorni za stvaranje inicijalnog
defekta na tubulima (2,4,5). Kada je barijera krv-testis jednom
narusena, antigeni i imuni kompleksi lako prolaze kroz peritubularnu bazalnu
membranu. T celijski odgovor koji se u ovom slucaju razvija biva potenciran
novim prodorima kroz barijeru krv-testis i zapaljenjskom reakcijom
koja zahvata citav organ.
Orhitis je
najcesca komplikacija mumpsa kod muskaraca u postpubertalnom periodu. Ovaj
oblik orhitisa se karakterise padom nivoa serumskog testosterona i
smanjenjem osetljivosti Leydigovih celija za gonadotropine (42). Detalji
patogenentskog mehanizma orhitisa posle mumpsa jos uvek nisu poznati, ali se
pretpostavlja da su antigeni parotidne zlezde slicni testikularnim
antigenima, sto dovodi do unakrsne reakcije antiparotidnih antitela i T
limfocita sa antigenima testisa. Kao rezultat ove unakrsne reaktivnosti
razvija se autoimunski orhitis (usmeno saopstenje). Neki autori smatraju da
je razvijanje autoimunskog orhitisa kao posledice infekcije virusom mumpsa
posledica aktivacije T celijskog imuniteta, povecanja serumske i tkivne
koncentracije proinflamatornih citokina i aktivacije makrofagalne funkcije u
samom tkivu testisa (43). Protivnici ove pretpostavke nalaze svoje
protivargumente u zapazanju da vecina drugih virusnih infekcija nije pracena
znacajnijom stopom komplikacija u vidu autoimunskog orhitisa.
Poaetite nasu stranicu
Spermogram
Animacije normalnog oplodjenja: Literatura:
1. Steco D.M.
et al. Regulation of the acute production of steroids in steroidogenetic
cells. Endocr Rev 1996; 17(3):244.


2. Blackwell T.S. et al. Sepsis and cytokines: current status. Br J Anesth
1996; 77:110.
3. Bosmann H.B. et al. Acute in vivo inhibition of testosterone by endotoxin
parallels loss of steroidogenic acute regulatory (StAR) protein in Leydig
cells. Endocrinology 1996; 137(10): 4522.
4. Head J.R. et al. Immune privileged in the testis. II: Evaluation of
potential local factors. Transplantation 1985; 40(3):269.
5. Hutson J.C. Testicular macrophages. Int Rev Cytol 1994; 149:99.
6. Hales D.B. Leydig cell-macrophage interactions: an overview. In: Payne
A.H. et al. The Leydig Cells. Viena, IL: Cache River Press, 1996:451.
7. Bergh A. Treatment with hCG increases the size of Leydig cells and
testicular macrophages in unilateral cryptorchid rats. Int J Androl 1987;
10:765.
8. Xiong Y. et al. Expression of TNF-a and IL-1b in mouse Leydig cells. Biol
Reprod 1993; 48(1):376.
9. Hutson J.C. Comparison of cellular and secreted proteins of macrophages
from the testis and peritoneum on two-dimensional polyacrylamide gels:
Evidence of tissue specific function. Reg Immunol 1989; 2:249.
10. Yee J.B. Bichemical consequences of follicle-stimulating hormone binding
to testicular macrophages in culture. Biol Reprod 1985; 32:872.
11. Sainio-Pollanen S. et al., CD28-CD80/CD86 interactions in testicular
immunoregulation. J Reprod Immunol 1996; 31(3):145.
12. Saz J.M. Leydig cells: Endocrine, paracrine and autocrine regulation.
Endocr Rev 1994; 15(5): 574.
13. Calkins J.H. et al. IL-1 inhibits Leydig cells steroidogenesis in
primary culture. Endocrinology 1988; 123:1605.
14. Hales D.B. IL-1 inhibits Leydig cells steroidogenesis primarily by
decreasing 17a-hydroxilase/C17-20 lyase cytohrome P450 expression.
Endocrinology 1992; 131(5):2165.
15. Xiong Y. et al. The role of TNF-a in the regulation of mouse Leydig
cells steroidogenesis. Endocrinology 1993; 132(6): 2438.
16. Guo H. et al. IL-2 is a potent inhibitor of Leydig cell steroidogenesis.
Endocrinology 1990; 127:1234.
17. Meikle A.W. et al. Reduction of testosterone synthesis after high dose
IL-2 therapy of metastatic cancer. J Clin Endocrinol Metab 1991; 73(5):931.
18. Kasahara T. et al. IL-2 mediate immune IFN-g production by human T cells
and T cell subsets. J Immunol 1983; 130(4):1784.
19. Orava M. et al. Human leukocyte IFN inhibits hCG stimulated testosterone
production by porcine Leydig cells in culture. Biochem Biophy Res Commun
1985; 127(3):809.
20. Orava M. et al. Treatment with preparations of human leukocyte
interferon decreases serum testosterone concentrations in men. Int J Cancer
1986; 38:295.
21. Saez J.M. Regulation of Leydig cell function by hormones and growth
factors other than LH and IGF-1. In: Payne A.H. et al. The Leydig Cell.
Vienna, IL: Cache River Press, 1996:383.
22. Ahmed S.A. et al. Sex hormones, immune responses, and autoimmune
diseases. Mechanisms of sex hormone action. Am J Pathol 1985; 121:531.
23. Cohn D.A. High sensitivity to androgen as a contributing factor in sex
differences in the immune response. Arthritis Rheum 1979; 22(11):1218.
24. Cutolo M. et al. Androgen replacement therapy in male patients with
rheumatoid arthritis. Arthritis Rheum 1991; 34(1):1.
25. Araneo B.A. et al. Dyhidrotestosterone exerts a depressive influence on
the production of IL-4, IL-5 and IFN, but not IL-2 by activated murine T
cells. Blood 1991; 78(3):688.
26. Sasson S. et al. Effect of androgenic steroids on rat thymus and
thymocytes in suspension. J Steroid Biochem 1981; 14:509.
27. Syed V. et al. Residual bodies activate Sertoly cell IL-1 release which
triggers IL-6 production by an autocrine mechanisms, through the
lypooxigenase pathway. Endocrinology 1995; 136:3070.
28. Hakovirta H. et al. Function of IL-6 as an inhibitor of meiotic DNA
synthesis in the rat seminiferous epithelium. Molec Cell Endocrinol 1995;
108:193.
29. De S.K. et al. Expression of TNF-a in mouse spermatogenic cells.
Endocrinology 1993; 133(1):389.
30. Pentikainen V. et al. TNF-a down-regulates the Fas ligand and inhibits
germ cell apoptosis in the human testis. J Clin Endocrinol Metab 2001;
86(9):4480.
31. Dejucq N. et al. IFN-a and IFN-g expression in the rat testis.
Endocrinology 1995; 136(11):4925.
32. Bussiere J.L. et al. Reproductive effects of chronic administration of
murine IFN-g. Reprod Toxicol 1996; 10(5):379.
33. Adekunle A.O. et al. Comparative analysis of blood plasma epidermal
growth factor concentrations, hormonal profiles and semen parameters of
fertile and infertile males. Afr J Med Med Sci 2000; 29(2):123.
34. Stuchi-Perez E.G. et al. Evaluation of the tubular and interstitial
functions of the testis in 46,XY patients with ambiguous genitalia. J
Pediatr Endocrinol Metab 2000; 13(6):605.
35. Gnessi L. et al. Leydig cell loss and spermatogenic arrest in
platelet-derived growth factor (PDGF)-A-deficient mice. J Cell Biol 2000;
149(5):1019.
36. Teuscher C. et al., Identification of 2 stallion sperm-specific proteins
and their autoantibody response. Equine Vet J 1994; 26(2):148.
37. Herr J.C. et al., Human antisperm monoclonal antibodies constructed
postvasectomy. Biol Reprod 1985; 32:695.
38. Lui W.Y. et al. Transforming growth factor-beta3 perturbs the inter-Sertoly
tight junction permeability barrier in vitro possibly mediated via its
effects on occludin, zonula occludens-1, and claudin-11. Endocrinology
2001; 142(5):1865.
39. Head J.R. et al., Immune privilege in the testis. I. Basic parameters of
allograft survival. Transplantation 1983; 36(4):423.
40. Kenneth W.B. Immunoglobulin Entry into the Male Reproductive Tract:
Implications for immunocontraception and vaccine development. In:
Reproductive Immunology Eds. Gupta S.K. Kluwer Academic Publishers, Boston,
1999: 285.
41. Bozas S.E. et al. Several vascular complement inhibitors are present on
human sperm. Biol Reprod 1993; 48(3):503.
42. Adamopoulos D.A. et al. Pituitary-testicular interrelationship in mumps
orchitis and other viral infections. Br Med J 1978; 1:1177.
43. Borden C.E. Interferons: pleotropic cellular modulators. Clin Immunol
Immunopathol 1992; 62(1):18.
{kind=link}
{kind=link}
{kind=link}